
제23강 뇌와 초월의식
1.The higher nervous system arrangements inhibit (or control) the lower, and thus, when the higher are suddenly rendered functionless, the lower rise in activity.
(예컨데, 전전두엽이 대상회나 시상이나 시상하부를 제어한다.) - John Hughlings Jackson (영국의 신경학자)
2.폴 맥클린의 뇌삼위일체설(Brain triune)
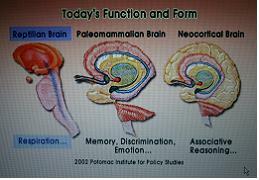 Reptilian brain(파충류적인 뇌) - 뇌간을 중심으로 발달, 호흡, 맥박 등과 관련
Reptilian brain(파충류적인 뇌) - 뇌간을 중심으로 발달, 호흡, 맥박 등과 관련
Paleomammalian brain - 고포유동물의 뇌, 감정을 generation하는 부위, 변연계
Neocortical brain - 신피질 발달
미국의 심리학자 폴 맥클린이 제창한 뇌 모형으로서 뇌삼위일체설이란 인간의 뇌를 쉽게 식별 가능한 세 개의 영역으로 나눌 수 있다는 것이다. 세 개의 영역이란 뇌간, 변연계, 대뇌피질을 말한다.
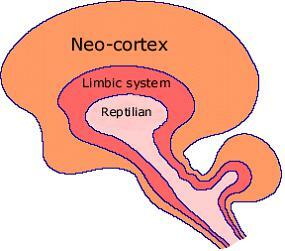 첫번째 영역인 뇌간(brainstem)은 가장 원시적인 부위이다. 뇌간은 뇌의 중심부로 뇌는 뇌간을 통해 척수와 결합된다. 파충류는 뇌간이 크고 뚜렸하기 때문에 매클린은 뇌간을 파충류의 뇌 또는 R 복합체(Reptile Complex)라고 불렀다.
첫번째 영역인 뇌간(brainstem)은 가장 원시적인 부위이다. 뇌간은 뇌의 중심부로 뇌는 뇌간을 통해 척수와 결합된다. 파충류는 뇌간이 크고 뚜렸하기 때문에 매클린은 뇌간을 파충류의 뇌 또는 R 복합체(Reptile Complex)라고 불렀다.
매클린의 뇌삼위일체론의 두번째에 해당하는 부위가 변연계 (limbic system) 이다. 변연계는 해마(hippocampus), 편도체(amygdala), 시상하부 hypothalamus) 등으로 구성되며 동물적 충동의 발현에 상황을 부여하는 역할을 담당한다.
변연계는 모양과 크기가 서로 다른 각 부분이 집합체를 형성하고 서로 연락을 주고 받는 복합 영역으로 그 부분들이 손상되면 이에 대응하여 기묘한 정서 반응이 일어난다. 예를 들어 변연계의 일부인 중격을 손상 당한 쥐는 아무런 이유없이 흉포해진다. 변연계의 또 다른 일부인 편도체를 손상 당한 인간은 클루버-버시 증후군(Kluver-Bucy syndrome) 이라는 기묘한 증상을 보인다. 클루버-버시 증후군 환자는 구애 행동과 성욕을 과도하게 드러낸다. 그들은 아무 물건이나 끊임없이 입으로 가져갈 뿐 아니라 놀랍게도 가구와 같은 무생물에 추파를 던지는 등 무분별하고 난잡하게 자신의 성욕을 드러내기도 한다. 그런 행동이 정서장애인 것은 분명하지만 감정 그 자체는 정상이다. 그릇된 것은 감정의 내용인 것이다.
뇌삼위일체론의 세 번째 영역에 해당하는 대뇌피질 (cortex) 은 논리와 선악 판단 등을 담당한다. 즉 대뇌 피질은 뇌간과 변연계에서 분출하는 근원적 행동을 억제하는 기능을 담당하는 것으로 보인다.
결론적으로 뇌삼위일체론이란 뇌의 기능을 단순화시켜서 뇌간은 본능을, 변연계는 감정을, 대뇌 피질은 지성을 담당한다고 정의하고 있다.
3.뇌의 hierarchy
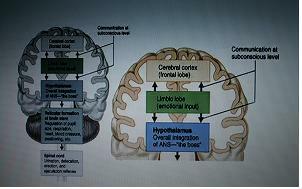 Cerebeal cortex - 특히 배외측전전두엽, 종합적인 컨트롤
Cerebeal cortex - 특히 배외측전전두엽, 종합적인 컨트롤
Limbic lobe - emotional input
Hypothalamus - overall integration of ANS(자율신경계의 관제탑)

Reticular formation of brainstem
(regulation of pupil size, respiration, heart, blood pressure, swallowing, etc)
Spinal cord
(urination, defecation, erection and ejaculation reflexes)
감정이나 느낌 뿐 아니라 종교적 체험, 초월의식을 설명하는데 변연계와 시상하부가 중요

뇌하수체는 brain속의 brain

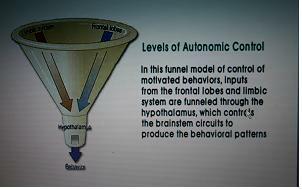
funnel model(깔대기 모델) : 대뇌피질과 변연계의 정보가 시상하부로 모여서 행위로 표출
4.시상하부(Hypothalamus)
시상하부는 간뇌(diencephalon)에 속하는 부분으로 전체 뇌중량의 0.5%가 채 안될 정도로 그 크기는 매우 작지만, 외부 또는 내부 환경의 변화에 따른 신경기전의 조절을 하는 매우 중요한 역할을 한다.
시상하부는 몸의 항상성(homeostasis)을 유지하기 위한 조절 중 내분비성 조절을 통제하는 신경내분비기관(neuroendocrine organ)이며, 자체에서도 호르몬을 생산하여 내분비작용을 한다. 또한 내장의 분비작용이나 평활근의 수축을 조절하는 자율신경계(ANS)의 중추가 되며, 외부 환경의 변화에 능동적으로 대처하는 여러 행동의 조절, 즉 식이(feeding behavior)의 조절, 성적행동(sexual behavior)의 조절 및 공격적 행동(aggressive behavior)의 조절 등 생명을 유지하고 생활에 적응하는데 필요한 여러가지 기능을 한다.
시상하부의 배쪽 경계는 비교적 뚜렷하여 앞쪽은 시각신경교차(optic chiasm)가 경계가 되고 외측은 시각로(optic tract)로 경계지워지며, 뒤쪽으로는 유두체(mammillary body)가 후방의 경계가 된다.
등쪽 경계는 불명확하며 등쪽 내측으로는 시상하고랑(hypothalamic sulcus)에 의해 시상과 구분되고 앞쪽은 대체로 제3뇌실의 앞쪽벽인 종말판(lamina terminalis)과 전교련(anterior commissure)에까지 이른다. 전교련의 아래쪽으로는 시각전구역(preoptic area)이 있으며 이 부분은 시상하부에 포함시키는 것이 일반적이다. 그 앞에는 중격핵(septal nuclei)과 브로카대각선조핵(nucleus of diagonal band of Broca)으로 이어진다. 후방으로는 중뇌의 중뇌수도관주위회색질(periaqueductal gray)과 피개(tegmentum)로 명확한 경계없이 이어지며, 외측 뒤쪽으로는 시상밑부(subthalamus)가 위치한다.
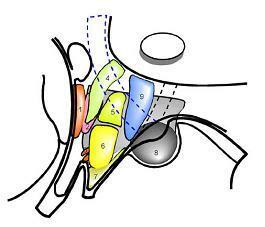
1.시각교차전구역(preoptic region)
2.전시상하부핵(anterior nucleus)
3.시각교차상핵(supraoptic nucleus) - 밤낮의 주기 조절
4.뇌실옆핵(paraventricular nucleus)
5.등쪽내측시상하부핵(dorsomedial nucleus)
6.배쪽내측시상하부핵(ventromedial nucleus)
7.깔대기핵(infundibular nucleus, 궁상핵 arcuate nucleus)
8.유두체핵(mammillary nucleus) 9.후시상하부핵(posterior nucleus)
그림 10-10. 시상하부(hypothalamus)의 핵군
시상하부는 세로방향으로 내측구역(medial zone)과 외측구역(lateral zone)의 두 구역으로 크게 나눌 수 있으며 이 중 내측구역은 상당히 넓고 뚜렷하여 다시 시각전구역(preoptic region)과 전시상하부구역(anterior hypothalamic region, supraoptic or suprachiasmatic region), 중간시상하부구역(intermediate hypothalamic region, tuberal region), 그리고 후시상하부구역(posterior hypothalamic region, mammillary region)으로 나눌 수 있다.
시각전구역(preoptic region)은 종뇌(telencephalon)에서 파생된 구조로 여겨지고 있지만 그 연결과 기능은 시상하부와 거의 같기 때문에 시상하부의 일부로 분류한다. 이 핵군은 제3뇌실의 앞쪽 시각전오목(preoptic recess)의 벽을 이루고 있는 뚜렷하지 않은 시각전뇌실옆핵(preoptic periventricular nucleus)과 그 바로 외측의 뚜렷한 내측시각전핵(medial preoptic nucleus), 그리고 외측시상하부핵의 앞에 위치한 외측시각전핵(lateral preoptic nucleus)으로 구성되어 있다. 이 부위의 신경원에서는 뇌하수체 전엽에서 분비되는 성선자극호르몬(gonadotropin)의 분비를 촉진하는 성선자극호르몬분비촉진호르몬(gonadotropin releasing hormone, GnRH)이 분비된다. 항상 일정한 농도로 분비되는 남성의 시각전구역이 월경주기에 따라 분비되는 여성의 시각전구역에 비해 더 커 성적으로 형태가 다른 핵(sexually dimorphic nucleus)으로 알려져 있다.
전시상하부구역(anterior hypothalamic region) 또는 시각상구역(supraoptic region, 시각교차상구역 suprachiasmatic region)은 전시상하부핵(anterior nucleus), 시각상핵(supraoptic nucleus), 뇌실옆핵(paraventricular nucleus, 실방핵)으로 구성되어 있다. 이 중에서 시각상핵과 뇌실옆핵은 뇌하수체 후엽에서 분비되는 호르몬인 항이뇨호르몬(antidiuretic hormone, ADH)과 자궁수축호르몬(oxytocin)을 분비하는 내분비성 핵이다.
중간시상하부구역(intermediate hypothalamic region) 또는 융기구역(tuberal region)은 등쪽내측시상하부핵(dorsomedial nucleus), 배쪽내측시상하부핵(ventromedial nucleus)과 깔대기핵(infundibular nucleus, 궁상핵 arcuate nucleus 베타엔돌핀이 분비되어 전전두엽으로 바로 정보를 보낸다)으로 구성되어 있다. 이 부분에서는 뇌하수체 전엽으로 시상하부-뇌하수체문맥계(hypothalamohypophyseal portal system)를 통해 여러가지 분비촉진 또는 분비억제호르몬을 방출하여 선뇌하수체에서 분비되는 여러 호르몬의 양을 조절하는 역할을 한다.
후시상하부구역(posterior hypothalamic region) 또는 유두체구역(mammillary region)은 내측유두체핵(medial mammillary nucleus), 중간유두체핵(intermediate mammillary nucleus), 외측유두체핵(lateral mammillary nucleus) 등 유두체에 위치하는 핵과 후시상하부핵(posterior nucleus)으로 구성되어 있다. 유두체는 해마형성체(hippocampal formation)에서 기원되는 뇌궁(fornix)의 섬유가 종지하는 곳이며 유두시상로(mammillothalamic tract)를 통해 시상과 연결되고 이는 다시 대상고랑(cingulate gyrus)을 거쳐 해마형성체로 연결되는 파페츠회로(Papez circuit)를 구성하는 부분이다.
외측구역(lateral zone)은 앞쪽과 뒤쪽은 좁고 중간은 넓으며 내측구역과는 유두시상다발(mammillothalamic tract)과 뇌궁기둥(column of fornix)에 의해 구분된다. 신경원은 세로로 주행하는 신경섬유다발 사이에 흩어져 있으며 외측시상하부핵(lateral hypothalamic nucleus)이라고 한다. 회색융기(tuber cinereum)의 심부, 내측구역 융기부의 외측에는 비교적 뚜렷하게 경계지워지는 두세 개의 외측융기핵(lateral tuberal nucleus)이 위치해 있다.
연결(connections)
시상하부는 변연계에 속하는 여러 구조와 복잡하게 서로 연결되어 있으며 뇌간의 그물형성체에 속하는 구조들과도 복잡하게 서로 연결되어 있다. 시상하부의 대표적인 전도로인 내측전뇌다발(medial forebrain bundle)과 등쪽세로다발(dorsal longitudinal fasciculus) 등은 대부분 구심성분과 원심성분이 함께 있는 복합전도로이며 그 기원부위와 종지부위도 중추신경계의 전역에 걸쳐 있다.
특히 중격-시상하부-중뇌 연속체(septo-hypothalamo-midbrain continuum)에 속하는 중격핵과 중뇌의 그물형성체와는 서로 고도로 연결되어 있다.
구심성연결(afferent connection): 시상하부로 들어오는 구심섬유는 (1) 직접 또는 뇌간의 그물형성체를 거쳐 내장구심성분을 전달하는 상행내장섬유(ascending visceral fibers)와 (2) 대뇌피질 중 계통발생학적으로 오래된 부분인 일차후각피질(primary olfactory area)을 포함하는 구피질(paleocortex)과 편도핵복합체(amygdaloid nuclear complex)에서 들어오는 성분, (3) 원시피질(archicortex)에 속하는 해마형성체(hippocampal formation)에서 기원되는 구심섬유로 나누어 볼 수 있다.
상행내장섬유(ascending visceral fibers)는 특수내장감각(SVA)인 미각(taste)과 일반내장감각(GVA)을 중개하는 고립로핵(solitary tract nucleus)과 부완핵(parabrachial nucleus) 등의 구조에서 부분적으로는 등쪽세로다발(dorsal longitudinal fasciculus, DLF)을 거쳐 그물형성체를 통해 시상하부의 여러 부위로 연결된다. 그물형성체의 솔기핵(raphe nuclei)과 청색반점핵(locus ceruleus) 등에서도 시상하부로 연결되며, 중뇌그물형성체의 등쪽 및 배쪽피개핵(dorsal and ventral tegmental nucleus)에서는 유두체각(mammillary peduncle)을 통해 외측유두체핵(lateral mammillary nucleus)으로 신경섬유를 보낸다.
일차후각피질에서는 내측전뇌다발(medial forebrain bundle)을 통해, 편도핵복합체(amygdaloid nuclear complex)에서는 분계선조(stria terminalis)와 배쪽편도원심성로(ventral amygdalofugal pathway)를 통해 시상하부로 원심섬유를 보낸다.
해마형성체(hippocampal formation)에서는 뇌궁(fornix) 섬유가 유두체(mammillary body)를 포함한 후시상하부구역(posterior hypothalamic area)으로 이어진다.
원심성연결(efferent connection): 시상하부에서는 뇌간과 척수의 자율신경신경절이전신경원(autonomic preganglionic neuron)으로 등쪽세로다발(dorsal longitudinal fasciculus, DLF)과 내측전뇌다발(medial forebrain bundle, MFB)을 통해 하행섬유를 보내며, 유두체(mammillary body)에서는 유두시상다발(mammillothalamic tract)을 통해 시상의 전핵군(anterior nuclear group)과 외측등쪽핵(lateral dorsal nucleus, LD)으로 상행섬유를 보내고, 일부 곁가지는 유두피개다발(mammillotegmental tract)을 통해 하행하여 등쪽피개핵(dorsal tegmental nucleus)과 교뇌그물핵(pontine reticular nucleus) 등 뇌간 상부의 그물형성체에 종지한다.
해마에서의 정보 유출은 뇌궁을 통해, 편도체에서 정보 유출은 분계선조를 통해서
모든 감각입력과 기억이 전전두엽으로 오고, 본능을 컨트롤하는 시상하부에서도 전전두엽으로 바로 정보를 보낸다.
5.변연계

1. 대상이랑(cingulate gyrus, 대상회)
2. 해마옆이랑(parahippocampal gyrus)
3. 해마형성체(hippocampal formation)
4. 이상엽(piriform lobe)
5. 중격부(septal region)
6. 편도체(amygdaloid body)
7. 시상하부(hypothalamus)
8. 시상(thalamus)
9. 시상상부(epithalamus)
10. 중뇌변연구역(midbrain limbic area)
그림 10-1. 변연계(limbic system)에 속하는 구조
오랫동안 뇌는 차가운 이성과 논리를 담당하는 기관으로 여겨져 왔고 정열은 심장, 식욕은 위, 성욕은 생식기 등 감정은 다른 기관의 기능으로 생각되어져 왔다. 그러나 이성과 논리와 마찬가지로 정열과 욕망, 환희와 열락, 분노와 비탄도 뇌의 기능적인 산물이다.
뇌에서 감정(emotion)을 담당하는 부위는 1937년 파페츠(Papez)가 대뇌 외측뇌실(lateral ventricle) 주위에 있는 피질구조-1878년에 프랑스의 해부학자인브로카(Broca)가 대변연엽(la grande lobe limbique)이라고 부른 구조-와 시상하부(hypothalamus), 그리고 시상(thalamus)을 연결하는 회로("파페츠회로 Papez circuit")에 있다고 주장한 이 후, 이들 구조들과 서로 밀접하게 연결되어 있는 편도체(amygdaloid body), 중격부(septal region), 시상하부(hypothalamus) 등과 함께 변연계(limbic system)라고 불리워진다.
외측뇌실을 둘러싸고 있는 변연엽(limbic lobe)에 속하는 구조는 두 개의 고리모양의 구조로 나눌 수 있다. 바깥쪽에는 궁상이랑(gyrus fornicatus) 즉, 대상이랑(cingulate gyrus)과 해마옆이랑(parahippocampal gyrus), 그리고 이 두 구조를 이어주는 대상이랑협부(isthmus)가 있으며, 안쪽에는 해마형성체(hippocampal formation), 즉 고유해마 (hippocampus proper), 치아이랑(dentate gyrus), 해마이행부(subiculum)와 이와 이어지는 작은 흔적구조(rudimentary structure)인 작은다발이랑(fasciolar gyrus, 소대회), 회색층(indusium griseum, 뇌량상이랑 supracallosal gyrus), 종말판옆이랑(paraterminal gyrus, 뇌량밑이랑 subcallosal gyrus), 뇌량밑구역(subcallosal area, 후각 옆구역 parolfactory area)이 여기에 속한다.종말판옆이랑(paraterminal gyrus)의 피질하에 있는 구조로 중격핵(septal nucleus)이 있다. 중격핵은 뒤쪽으로는 외측뇌실 사이에 있는 투명중격(septum pellucidum)으로 이어진다. 측두엽(temporal lobe) 전방의 피질 아래에는 회색질 구조인 편도체(amygdaloid body), 즉 편도핵복합체(amygdaloid nuclear complex)가 있다. 이들 구조들은 모두 시상하부(hypothalamus)와 연결되어 있으며 시상하부의 여러 부분을 전기적으로 자극하였을 때 기쁨, 슬픔, 분노, 식욕 등의 감정적 반응이 일어나는 것을 보아 이 부분이 감정의 표현과 밀접한 관계가 있다고 생각된다. 이와같은 감정적 반응이 일어나는 곳은 시상하부와 보통 시상하부의 일부로 간주되는 시각전구역(preoptic region), 그리고 중격핵 부위이며, 시상하부는 중뇌의 그물형성체(midbrain reticular formation)와 고도로 연결되어 있기 때문에 이를 중격-시각전구역-시상하부-중뇌 연속체(septo-preoptico-hypothalamo-midbrain continuum)라고도 한다. 시상하부와 연결된 중뇌의 그물 형성체에는 배쪽피개구역(ventral tegmental area, VTA), 등쪽피개핵(dorsal tegmental nucleus of Gudden), 등쪽솔기핵(dorsal raphe nucleus), 정중솔기핵(median raphe nucleus 상중심핵 superior central nucleus of Bechterew), 중뇌수도관주위회색질(periaqueductal gray) 등이 속하며, 이 부분을 중뇌변연구역(midbrain limbic area)이라고 한다.
변연계에 속하는 구조 중에는 일차감각으로 후각(olfaction)을 받아들이는 부위들이 있으며, 이러한 부위를 후각뇌(rhinencephalon)라고도 한다. 후각뇌는 넓은 의미로는 변연계를 포함하여 후각에 의해 영향을 받는 대뇌의 모든 부위를 가리키는 용어로 사용되기도 하나, 일반적으로는 후각망울(olfactory bulb)과 후각로(olfactory tract) 등 후각신경(olfactory nerve, I)을 통해 후각을 직접 받아들이는 부위와 후각망울에서 나오는 원심성 신경섬유가 직접 연결되어 있는 부분, 즉 이상엽(piriform or pyriform lobe)과 중격부(septal region) 및 편도체(amygdaloid body)의 피질내측핵군(corticomedial nuclear group)을 포함하는 구조를 말한다. 이상엽은 전이상엽피질(prepiriform cortex), 편도체주위피질(periamygdaloid cortex), 내후각뇌피질(entorhinal cortex)로 구성되어 있다.
시상하부 이외에도 간뇌(diencephalon)의 구조 중에는 변연계의 나머지 부분과 고도로 연결된 구조가 있다. 시상(thalamus)의 시상전핵군(anterior nuclear group)과 외측등쪽핵(lateral dorsal nucleus, LD)은 시상하부와 변연엽의 대상이랑(cingulate gyrus)을 이어주는 중요한 중계핵이며, 시상내측핵군(medial nuclear group)에 속하는 등쪽내측핵(dorsomedian nucleus)과 시상중간핵군(midline nuclei)도 이러한 구조들과 밀접하게 연결되어 있어 변연계의 일부로 분류한다. 시상상부(epithalamus)에 속하는 고삐핵(habenular nuclei) 역시 시상하부 및 중뇌변연구역과 고도로 연결된 변연계의 구조이다.
전전두엽피질(prefrontal cortex)과 측두엽 연합피질(temporal association cortex)도 변연계에 속하는 구조와 고도로 연결되어 있어 변연계의 일부로 포함시키기도 한다.
6.자율신경계
자율신경계(autonomic nervous system)는 체신경계(somatic nervous system)와 반대되는 용어로, 자신의 의지대로 제어할 수 없는 심장혈관계, 호흡기계, 소화기계, 비뇨생식기계 등 내장(viscera)의 활동과 관련된 신경조직을 말한다. 말초신경계에서 자율신경계는 교감신경계(sympathetic nervous system)와 부교감신경계(parasympathetic nervous system)의 두 부분으로 나누어진다. 이 두 신경계는 자율신경계의 원심성분이며, 대체로 길항적으로 작용한다. 교감신경계는 위급한 경우에 빠르게 대처할 수 있게(싸우거나 도망가는 반응 fight or flight reaction) 도와주는 역할을 한다. 교감신경계가 활성화되면 골격근과 신경계에 혈액공급이 늘어나며 심장의 박동이 증가하고 감각이 예민해진다. 반면에 내장으로 가는 혈액공급은 크게 감소한다.
부교감신경계는 이러한 위급한 경우를 대비해서 미리 에너지를 비축해 두는 역할을 한다. 내장으로 가는 혈액공급이 증가되어 소화, 배설 등이 활성화되고, 심장의 박동이 느려져 전체적인 에너지의 사용이 감소하며, 생식기관은 활성화된다.
교감신경계와 부교감신경계는 모두 두 개의 신경원 사슬로 구성되어 있다. 첫번째 신경원은 중추신경계 내에 있으며, 두번째 신경원은 말초신경계의 자율신경절(autonomic ganglion)에 위치해 있다. 첫번째 신경원의 축삭으로 구성된 신경절이전섬유(preganglionic fiber)는 두번째 신경원과 이어져 있고, 두번째 신경원에서 나오는 신경절이후섬유(postganglionic fiber)는 각 장기의 효과기와 연결된다. 자율신경계의 말초자율신경계에서 중추신경계로 이어지는 구심성분도 있으며, 그 수는 원심섬유보다 훨씬 많다.
자율신경계에 속하는 다른 한 부분으로 장신경계(enteric nervous system)가 있다. 이 계통은 소화관의 벽 내부에 위치해 있는 신경원들(장신경절 enteric ganglion)로 구성되어 있으며, 장에 미치는 장력과 화학적 환경의 변화에 따라 장과 혈관의 평활근의 수축도를 조절하고, 장분비세포의 분비활동을 변화시켜, 항상성(homeostasis)을 유지해주는 역할을 한다. 장신경계는 교감신경계와 부교감신경계의 영향도 받는다. 자율신경계는 말초신경계에만 있는 것이 아니라 중추신경계에도 존재한다
그림 16-8. 부교감 지배의 분포(Distribution of Parasympathetic Innervation).

그림 16-5. 교감부(The Sympathetic Division).
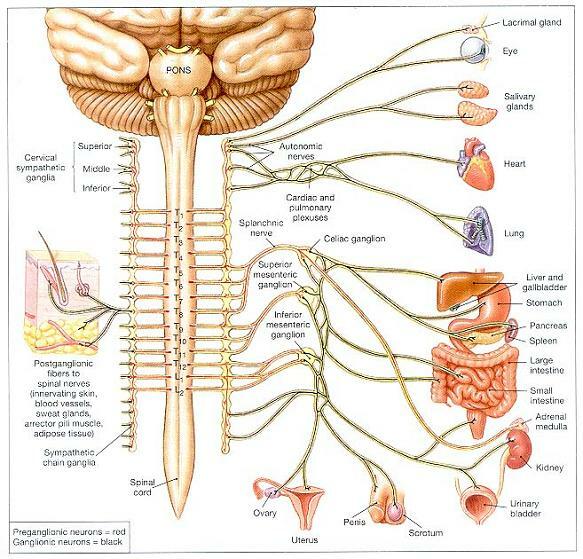
교감섬유의 분포는 양쪽이 동일하다. 그러나 명쾌하게 보이도록, 체성 구조물에 대한 지배는 왼쪽에, 내장성 구조물에 대한 지배는 오른쪽에 나타내었다.
7.종교적 초월현상(시상하부에 중점을 두고 접근)
능동적 명상 - 수피즘 / 수동적 명상 - 사마티
연합영역(초월의식과 관련깊은 것이 주의연합영역, 정위연합영역)
시각연합영역
주의연합영역(attention) - 전전두엽
정위연합영역(orientation) - 두정엽
언어개념연합영역
<초월상태의 뇌생리> (능동적 접근)

우 전전두엽(PFC, 주의연합영역에서 강력한 의지로 집중, 예컨대 화두, 염불 등)
→우 시상(그물핵, GABA 억제 신호) →우 두정엽(후상두정엽 PSPL, 정위영역) →해마 →편도체 →우 시상하부(복내측시상하부, 강력한 억제신호가 시상하부에서 강력한 흥분신호 발생), 우 시상하부에서 좌 시상하부로 강력한 breakthrough, 좌 시상하부(외측) →기저핵(Ach 시스템) → 좌 시상하부(궁상핵, 베타엔돌핀) → 좌 전전두엽 → 우 전전두엽 : 폐 loop를 그린다.
우 시상하부 →부교감신경
좌 시상하부 →좌 편도체 →해마 →좌 두정엽 →좌 시상 →좌 전전두엽
좌 시상하부 →교감신경
초월명상 시 정위영역에서의 변화
우측 - 수입로 차단 (우 두정엽은 3차원 공간을 지각)
좌측 _ 수입로 차단 (좌 두정엽은 팔이 뻗을 수 있는 범위내의 자극을 받아들인다 - 몸체에서 오는 자극을 process, 자기를 인식)
수동적 명상에서 좌우 두정엽의 수입로 자극이 차단
우측 - 공간(시간)이 사라짐 / 좌측 - 자아감이 사라짐 ⇒ 초월적 일체감, 물아일체(尋牛圖
의 일원상, 人牛俱忘 목동(self)도 소(바깥 경계)도 사라진 상태), 생각과 언어, 감각이 사라짐 → 순수한 인식 상태만 남는다 → 초월적 인식(자아가 없는 인식이 가능하다 : 諸法無我)
The Effect of Meditation on the Brain activity in Tibetan Meditators:
Frontal Lobes
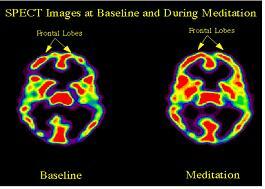
SPECT : single photon emission computed tomography
The first image (above) shows that the front part of the brain, which is usually involved in focusing attention and concentration, is more active during meditation.

The second image shows that there is decreased activity in the parietal lobe. This area of the brain is responsible for giving us a sense of our orientation in space and time. We hypothesized that blocking all sensory and cognitive input into this area during meditation results in the sense of no space and no time which is so often described in meditation.
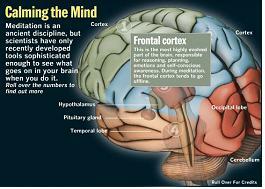
Frontal cortex :
This is the most highly evolved part of the brain, responsible for reasoning, planning, emotions and sel-conscious awareness. During meditation, the frontal cortex tends to go offline.
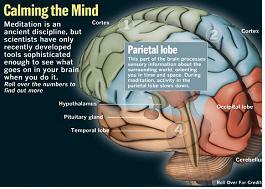 Parietal lobe :
Parietal lobe :
This part of the brain processes sensory information about the surrounding world, orienting you in time and space. During meditation, activity in the parietal lobe slows down.
초월적 상태에 들어가는 순간, 1/100초 만에 자극이 완전히 차단됨
 Thalamus :
Thalamus :
The gatekeeper for the senses, this organ focuses your attention by funneling some sensory data deeper into the brain and stopping other signals in their tracks. Meditation reduces the flow of incoming information to a trickle.
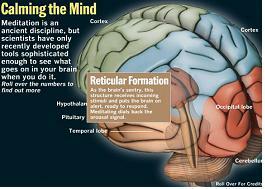
Reticular Formation :
As the brain's sentry, this structure receives incoming stimuli and puts the brain on alert, ready to respond. Meditation dials back the arousal signal.
신은 왜 우리 곁을 떠나지 않는가?
“신경신호의 완전한 차단은 오른쪽과 왼쪽 정위영역 모두에 극적인 효과를 미친다.
우리가 물리적 공간으로 경험하는 신경학적 토대를 만들어 내는 일을 담당하는 오른쪽 정위영역은 그 속에서 자신의 위치를 정할 수 있는 공간적 내용을 만드는데 필요한 정보를 결여한다.“
8.종교적 마음과 뇌와 실제적 관련 예
“1978년 여름 나는 주목을 끌어보려고 어리석은 허세를 부리느라 존 웨인의 레드리버 농장 침액 탱크에서 헤엄을 쳤다. 이 일로 내가 하는 일을 널리 홍보할 수 있었고, 몇 군데서 강연을 할 기회도 얻게 되었다. 그러나 유기인산 화합물이라는 화학물질이 몸에 묻어 나한테 끔찍한 영향을 미쳤다. 믿음에 대해 생각할 때 느꼈던 경외감이 갑자기 사라져 버린 것이다.”
“유기인산 화합물은 뇌안의 아세틸콜린 등 신경전달물질의 농도를 변화시키는 것으로 알려져 있다. 나는 또한 이 물질 때문에 생생하고 끔찍한 악몽에 시달렸다. 그렇지만 이 물질이 왜 종교적 외경심을 흔들어 놓았는지는 알 수가 없다. 모든 마법이 사라지고 실제 오즈의 마법사는 커튼 뒤에서 버튼이나 눌러 대는 왜소한 할아버지라는 사실을 알게 된 것과 비슷했다.”
“나는 또 신문에서 뉴욕 공공도서관 직원 한 사람이 지구상에서 영원불멸성이 존재하는 곳은 도서관 뿐이라고 말했다는 기사를 읽었다. 도서관은 인류의 축적된 기억이 존재하는 곳이다. 나는 이 말을 현판에 적어 책상 위에 걸어 놓았다. 그 말 덕에 끝까지 포기하지 않고 결국 박사 학위를 따낼 수 있었다.” 나는 그림으로 생각한다.
9.보조스님의 無心 공부 10가지
覺察(마음의 잡념에 생기는 순간 알아차리고) 休歇(놓아버려라, 방하착)
泯心存境(마음을 없애고 경계를 둔다) 泯境存心
泯心泯境 存心存境
內外全体 內外全用
卽体卽用 透出体用