제18강 운동성의 기원
1.사고는 운동성의 진화적 내면화이다(이나스)
의식과 사고를 궁극적으로 공부하려면 세포부터(특히 세포막, 이온채널(단백질))
미토콘드리아는 우리 몸의 모든 세포에 있다.
운동성을 추적하다보면 미토콘드리아에 귀착하게 되고, 또 그 쌍인 엽록체를 보게 되며 결국 ATP(모든 생명체의 통화량)를 알아야 한다.
모든 생명 시스템의 궁극적인 운동성, 에너지 흐름은 ATP로 귀결
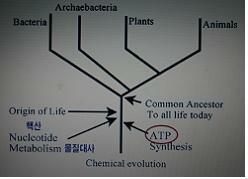
생명현상의 주역 : DNA와 ATP
“복제자(DNA)가 생명드라마의 주인공이라면 효소(ATP)는 조연이 아니라 공동 주연이다.”
-조상이야기(리처드 도킨스)
“다세포체란 한 종의 개체발생이 거치는 특정의 순간일 뿐이며, 그 종의 변이는 변함없이 세포수준에서 일어난다.“ -인식의 나무-
고향의 강
눈감으면 떠오르는 고향의 강 → DNA강 또는 양성자 강(빅뱅이후 찰라적 순간에 양성자 형성)
지금도 흘러가는 가슴 속의 강

“우리 관점에서 생물들을 보면, 당신네 진핵생물들은 곧 그런 젠체하는 태도를 버릴 것이다. 당신네 두발 유인원들, 꼬리 잘린 나무땃쥐, 물기가 빠진 육기어류, 척추를 가진 벌레들, 혹스유전자를 늘린 해면동물들, 한꺼번에 무더기로 등장한 것들, 진핵생물들, 단조롭고 비좁은 영역에 옹기종기 모인 거의 구별되지 않은 존재들은 세균들의 표면에 있는 변덕스러운 거품에 다름 아니다. 당신들을 이루는 세포들 자체가 우리 세균들이 10억년 전에 발견한 낡은 기술들을 똑같이 재현하는 세균 군체들이기 때문이다. 우리는 당신들이 오기 전부터 여기에 있었고, 당신들이 떠난 뒤에도 여기에 남을 것이다.” - 조상 이야기
2.광합성
가.엽록체(Chloroplast)
잎의 엽육조직에 있는 전형적인 식물세포는 50개 이상의 엽록체를 가지고 있다. 엽록체는 3종류의 막으로 이루어져 있다.
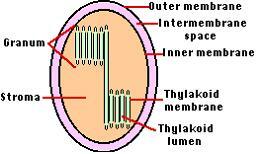
-외막 : 매끈하며 분자들이 자유롭게 통과한다.
-내막 : 매끈하며 많은 수송체(transporter)를 가진다. 수송체는 막단백질로 분자들의 출입을 조절한다.
-틸라코이드 막
1) 틸라코이드(Thylakoid)
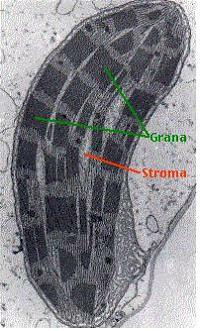
틸라코이드막은 내강(lumen)을 둘러싸는 일종의 소포계를 만든다. 이들 소포계는 거의가 서로 연결되어 있다. 엽록체에는 여러 곳에서 이들 소포체가 쌓여져 그라나(grana-동전더미모양)를 이룬다.
틸라코이드 막에는 4종류의 단백질이 존재하는데,
- 엽록소와 카로티노이드를 가지는 광계I
- 엽록소와 카로티노이드를 가지는 광계II
- 시토크롬 b와 f
- ATP합성효소(synthetase)
이들 단백질들은 소위 말하는 광합성의 명반응을 수행한다. 틸라코이드 막은 액체물질인 스트로마로 둘러 싸여 있다. 스트로마에는 광합성의 암반응을 수행하는데 필요한 RUBISCO와 같은 효소들이 존재한다.
2) 엽록소(Chlorophyll)
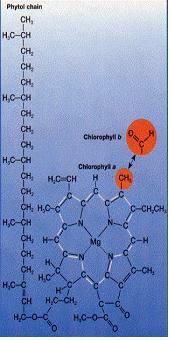
식물과 녹조류에서 2종류의 엽록소가 발견된다. 엽록소a, 엽록소b가 바로 그것이다. 이 둘의 구조 차이는 왼쪽 그림에서 붉은 원으로 표시되어 있다. 엽록체에서 두 종류의 엽록소는 틸라코이드 막에 내막단백질과 같이 존재한다. 포르피린 고리 주위에 교대로 단일결합과 이중결합이 존재한다. 이중결합에 있는 여분의 전자는 자유롭게 링 주위를 이동하고 있으며 이러한 분자적 특징에 의해 빛을 흡수할 수 있게 된다.
엽록소는 빛의 스펙트럼에서 적색과 자색부위를 가장 강하게 흡수한다. 녹색광은 거의 흡수되지 않고 반사되며 식물 잎의 색깔이 녹색으로 보이는 이유가 바로 여기에 있다.
3) 카로티노이드(Carotenoid)
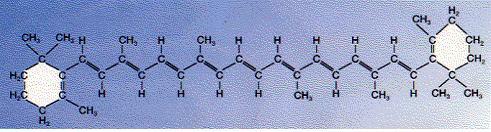
엽록체는 또한 카로티노이드도 가지고 있다. 이 화학분자들은 적색부터 황색까지의 색을 가지고 있다. 카로티노이드는 스펙트럼의 청색광을 더 강하게 흡수한다. 이들은 엽록소가 잘 흡수하지 못하는 빛을 흡수해주게 된다. 잘 익은 붉은 색 토마토와 당근의 오렌지 색은 바로 카로티노이드의 색이다.
잎에서 카로티노이드는 보통 엽록소에 의해 가려져 있다. 가을에 잎에 존재하는 엽록소의 양이 감소하면서 카로티노이드는 겉으로 드러나게 되어 가을철 단풍의 색깔이 나타나게 된다. 아래 그림은 가장 풍부한 베타-카로틴 구조를 보여 준다. 엽록소에서와 마찬가지로 이중결합과 단일결합이 교대로 존재하는 것을 볼 수 있다. 전체적인 구조는 두 개의 벤젠고리를 탄화수소 사슬이 연결해 주고 있는 모양이다. 많은 동물들은 비타민A의 전구물질로 베타-카로틴을 사용한다. 베타-카로틴을 정확히 이등분하면 바로 비타민A가 되며 벤젠고리에 붙어 있는 측쇄 중 일부가 산화되면 망막의 중요한 분자인 레티날이 만들어 진다.
나.PHOTOSYNTHESIS(광합성)
1) PHOTOSYNTHESIS : AN OVERVIEW
광합성(photosynthesis) : 태양에너지가 특정 분자들의 결합에 의해 포도당의 생성에 이용되는 과정. 반응물은 CO2와 H2O이며, 생성물은 포도당(C6H12O6), H2O ,O2이다.
6CO2 +12H2O → C6H12O6 + 6O2 + 6H2O(△G + 686kcal/mole ; 흡열반응)
포도당이 합성되는 과정에서 빛에너지가 ATP나 NADPH와 같은 에너지 물질의 합성에 이용
 1) 광합성의 두 부분
1) 광합성의 두 부분
① 명반응(light reaction) : 빛을 요구하는 반응이며, 계에 에너지를 충전시킴
태양에너지는 ADP와 NADP+를 각각 ATP와 NADPH로 전환시키는 데에 사용
12H2O + 18ADP + Pi + 12NADP+ → 18ATP +12NADPH + 12H+ + 6O2
※ 명반응의 생성물은 암반응의 반응물로 사용된다.
② 암반응(dark reaction) : 빛을 요구하지 않는 반응이며, 계를 원상태로 되돌림
ATP와 NADPH는 포도당 생성에 사용된 후 원래 상태로 되돌아감
6CO2 +18ATP + 12NADPH + 12H+ ->C6H12O6 +18ADP+Pi+12NADP+ + 6H2O
2) CHLOROPLASTS(엽록체)
1) 틸라코이드
그라나 틸라코이드와 스트로마 틸라코이드는 루멘(lumen)으로 알려진 통로로 연속된 내부공간을 갖으며, 루멘과 스트로마는 막에 의해 격리
① 두 광계(two phtosystem)
ㆍ틸라코이드막에 있는 빛을 포획하고 이용하는 특수구조로 광계I(photosystem I ; PS I)과
광계II(photosystem;PS II) 복합체가 있다.
ㆍ광수확안테나(light-harvesting antenna)와 반응중심(reaction center)을 갖으며, 전자전달계 와 밀접하게 연결
② 엽록소와 광수확 안테나
ㆍ광수확안테나 : 빛을 포획하여 에너지를 축적하는 일을 하며 엽록소a, 엽록소b (다른 영역 의 파장 흡수), 카로티노이드(carotenoid)로 구성
ㆍ반응중심(reaction center) : 에너지가 축적되는 지점으로 빛에너지가 화학에너지로 전환되는 부위. 한 분자의 엽록소a와 단백질로 구성. 엽록소a를 한 분자씩 갖고 있는 PS I과 PS II 의 반응중심은 각각 700nm 와 680nm 파장 흡수. 엽록소a가 안테나로부터 충분한 에너지를 흡수하면 엽록소의 전자가 들뜨게 되어 궤도에서 이탈 → 엽록소a에서 궤도를 이탈한 전자는 즉시 에너지를 방출하지 않고 전자전달계에 포획되어서 화학적인 일을 하는데 사용(광합성의 특징)
③ 전자전달계(electron transport system)
ㆍ빛에 의해 활성화된 전자의 에너지를 이용하여 양성자(proton)를 틸라코이드 루멘에 농축 → 자유에너지를 ATP 생성에 사용할 수 있게 함
전자가 NADP+를 NADPH+H+로 환원시키는데 이용
④ CF0CF1복합체 : ADP의 인산화에 의해 ATP 생성과정에 중요한 기능
ㆍCF0복합체 - 루멘에서 시작해 틸라코이드 막을 가로지르는 단백질에 의해 형성된 통로
ㆍCF1복합체 - 양성자의 농도기울기에 의해 CF1 머리 방향으로 양성자 이동
→ 농축된 양성자의 자유에너지를 이용해 ADP를 인산화(ATP 합성효소)
3) THE LIGHT REACTIONS OF PHOTOSYNTHESIS
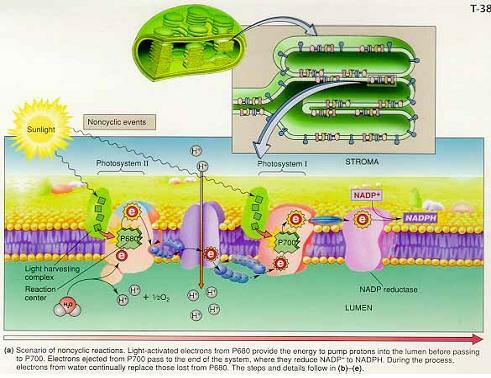
photosystem Ⅱ(PS Ⅱ) → plastoquinone(PQ) → 시토크롬bf → plastocyanin(PC) →photosystem I(PS I) → ferredoxine(FD) → NADP reductase
-비순환적 광인산화(noncyclic photophosphorylation) : PS I과 PS II가 함께 작용하여 화학삼투기울기를 형성하고 NADP+를 NADPH로 환원
-순환적 광인산화(cyclic photophosphorylation) : PS I만 작용하는 경우로 ATP 합성을 위한 화학삼투기울기만 형성
1) 비순환적 과정 : 빛이 PS II의 광수확안테나에 흡수됨으로써 시작
① 물과 P680의 환원
: 엽록소 a - 빛에너지에 이해 P680 반응중심의 엽록소a에선 전자 방출(산화)
산화된 엽록소a는 망간을 함유한 단백질에 의해 환원
→ 망간을 함유한 단백질은 유실된 전자를 루멘에 다량 존재하는 물로부터 얻음.
물분자 하나가 분해되면 두 개의 전자, 양성자 그리고 산소원자가 발생
H2O → 2e- + 2H+ + 1/2O2
물에서 하나의 전자가 분리될 때 동시에 하나의 양성자가 루멘으로 방출되어 양성자 기울기 형성을 돕는다.
⇒ 시아노박테리아가 물에서 전자를 탈취할수 있는 능력을 얻은 것이 생명 역사에서 가장 중요한 사건 중의 하나이다.
② 전자에너지와 양성자 수송
: P680 반응중심에서 방출된 들뜬 전자 : plastoquinone(PQ)으로 전달되어 스트로마의 양성자를 포획해 루멘으로 방출
→ 전자 운반자인 거대 시토크롬 복합체에 전달
→ 낮은 자유에너지 상태인 전자는 plastocyanin(PC)에 의해 PS I로 이동
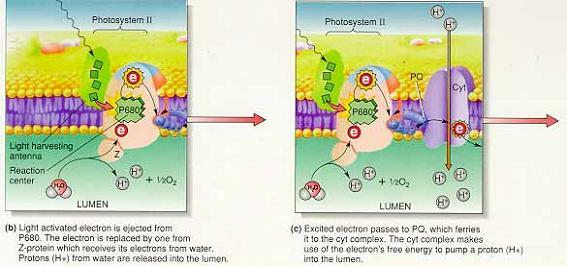
※양성자 농도기울기 형성 : PQ에 의해 스트로마로부터 유입되는 양성자와 물의 분해에 의해 생성되는 양성자에 의해 형성
③PS I : NADP+의 환원
-P700은 반응중심의 엽록소가 산화된 상태에서만 P680의 전자를 유입
(P700 반응중심의 환원은 P680 전자에 의해 일어남)
-PS I의 역할 : 들뜬 PS I 전자 에너지를 사용해 NADP+를 NADPH로 환원
(P700 반응중심에서 온 2개의 전자와 스트로마로부터 유입된 2개의 양성자가 필요)
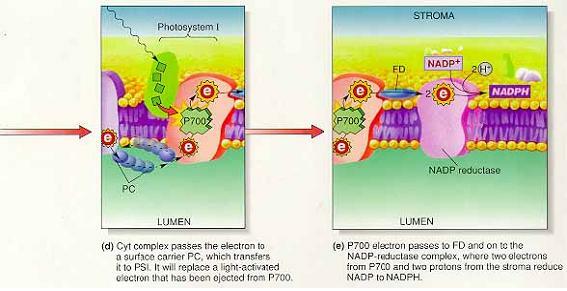
※ 비순환적 과정 요약
1. P680에 의한 광자 흡수
2. 들뜬 P680 전자가 전자전달계로 이동
3. P680 전자 재충전, 루멘에 양성자 증가, 기체 산소가 방출되는 물의 산화
4. P680 전자의 자유에너지를 이용하여 스트로마의 양성자를 루멘으로 유입
5. P700에 의한 광자 흡수
6. P680 전자가 P700 전자 대체
7. 들뜬 P700 전자의 전자전달계로 이동
8. NADP+가 NADPH로 환원될때 P700 전자의 에너지 이용
2) 순환적 과정
: NADPH를 생성하지 못하는 대신 P700 전자의 에너지를 이용해 스트로마로부터 루멘으로 양성자를 펌프한다. 따라서 ATP의 생성만을 돕는다. 암반응이 산화된 NADP+를 명반응으 로 되돌려 보낼때까지 계속...
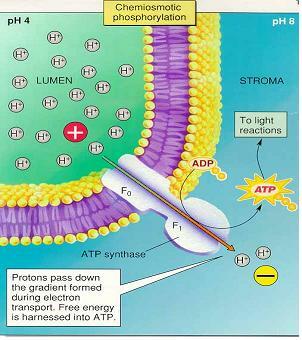
3) 화학삼투적 인산화
ㆍ양성자가 루멘에 축적되면 스트로마의 염소이온(Cl-)을 끌어 당겨 루멘에 HCl(염산)이 농축
ㆍ스트로마의 양성자가 루멘으로 방출되면 스트로마는 알칼리성 혹은 염기성을 띠게됨 → 화학삼투계는 농축된 산성용액(pH 4-5)이 되며 틸라코이드막에 의해 스트로마의 알칼리성용액(pH8)과 격리 ⇒ 양성자가 농도기울기에 의해 CF0 통로를 따라 ATP합성효소를 포함한 CF1 복합체에 도달하여 ATP 합성
4) THE LIGHT-INDEPENDENT REACTIONS
: 명반응을 통해 엽록체는 상당량의 퍼텐셜에너지를 저장하고 ATP와 NADPH 형태로 환원 력을 쌓아올리기 때문에 CO2만 유입되면 포도당 생성이 개시될 수 있음
6CO2 +18ATP + 12NADPH + 12H+ ->C6H12O6 +18ADP+Pi+12NADP+ + 6H2O
1) 캘빈회로(M. Calvin)
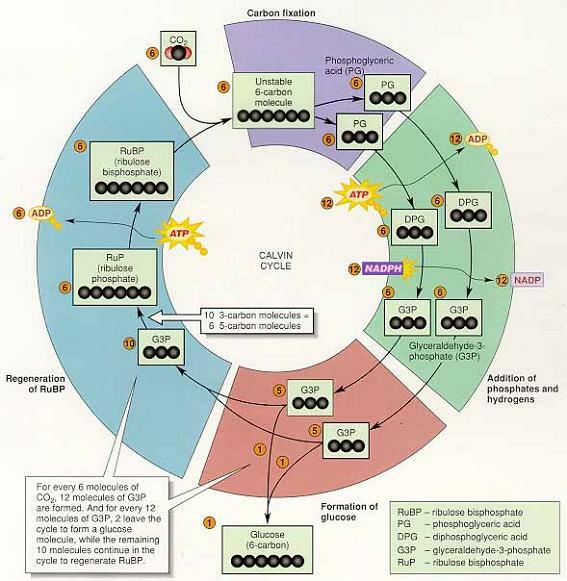
반응1. 카르복시화 반응(carboxylation) : CO2가 첨가되는 반응
리블로스이인산 카르복시화효소(ribulose-1,5-bisphosphate carboxylase, Rubisco) ⇒ CO2를 리블로스이인산(ribulose 1,5-bisphosphate, RuBp)에 결합시켜 불안정한 6탄당 중간대사물을 형성해 2개의 3탄당으로 분해
반응2. 인산화 : ATP의 고에너지 인산기가 2개의 3탄당 탄수화물로 전이(기질의 자유에너 지 증가)
반응3. 환원 : NADPH가 반응2.의 생성물이 갖고있는 인산기 중 하나를 수소로 치환, 떨어진 인산은 NADP+ 및 ADP와 함께 명반응으로 되돌아감
반응4. 반응3.의 생성물인 글리세르알데히드-3-인산(glyceraldehyde-3-phosphate,G-3-P)가 포도당(기타 생성물)을 생성하는 경로 또는 재생경로(regeneration pathway ; 캘빈회로 를 계속 유지)를 선택
① 포도당과 기타 생성물
포도당의 생성과정 : G-3-P + G-3-P (지방산, 글리세롤, 아미노산 합성에 이용,
↓ 미토콘드리아로 이동된 후 ATP 생성에도 이용)
과당-1,6-이인산fructose-1,6-disphosphate
↓
포도당-1-인산glucose-1-phosphate
↓
포도당 형성
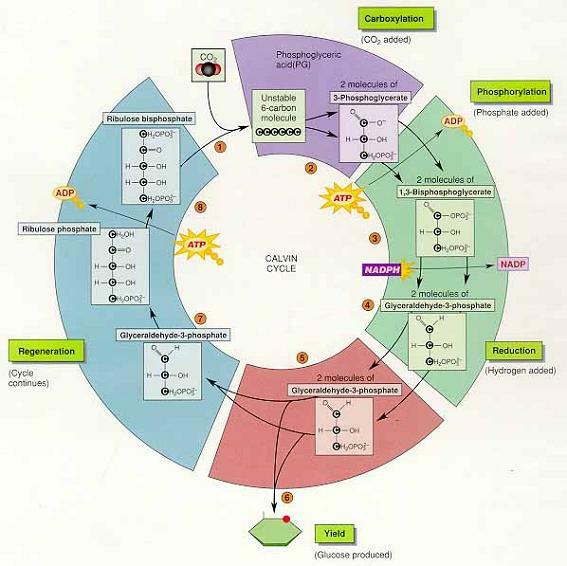
② G3P, 포도당, 캘빈회로의 유지
ㆍ캘빈회로가 5회 돌아 생성된 10분자의 G3P는 재생에 이용되고 6회 돌때 생성된 2분자 의 G3P는 포도당 형성에 이용
ㆍ한 분자의 CO2 가 투입되어 회로가 1회전할 때마다 2개의 NADPH, 2개의 H+와 3개의 ATP가 소모(포도당 한 분자를 합성하려면 캘빈회로가 6회전을 해야 하므로 6개의 CO2, 12개의 NADPH와 H+ 그리고 18개의 ATP가 필요)
6CO2 +18ATP + 12NADPH + 12H+ ->C6H12O6 +18ADP+Pi+12NADP+ + 6H2O
5) PHOTORESPIRATION : TROUBLE IN THE CALVIN CYCLE
캘빈회로를 정상적으로 작동시키는 Rubisco의 CO2포획능력은 CO2가 감소하거나 확산 속도보다 더 빠르게 고정되면 효소의 효율이 현저히 감소. CO2의 농도가 최소 수준 이하로 떨어지면 캘빈회로에서 O2가 CO2를 대체하여 광호흡 (photorespiration)을 함 ⇒ 산소와 리블로스 이인산이 결합하여 생성된 물질은 여러 단계를 거쳐 글리콜산(glycolate)이 됨
→ 세포질의 미소체(microbody)로 이동해 글리옥실산으로 전환
→ 미토콘드리아로 들어가 CO2와 3-PG로 전환 → CO2는 방출되고 3-PG는 연료로 사용
1) C4 탄소 고정
ㆍC4경로(M.D. Hatch & C.R. Slack) : 4탄소 화합물이 관여하는 새로운 CO2 고정 경로
ㆍC3식물 :탄소 고정이 캘빈회로에 의해서만 일어나고 카르복시화에 의한 첫번째 산물이 3탄소 화합물이다.
ㆍC4 식물 :CO2는 C3 식물에서와 같이 엽육세포(leaf mesophyll cell)의 엽록체로 확산
ㆍPEP카르복시화효소(phosphoenolpyruvate carboxylase)-CO2가 처음 만나는 효소로 매우 낮은 CO2 농도에서도 효율적으로 작용
ㆍ4탄소 대사경로의 최종 산물인 말산(malate)이 엽육세포로부터 인접한 유관속초 세포(bundle sheath cell, 엽맥을 에워싸고 있는 세포)의 엽록체로 이동 → 4탄소 기질은 3탄소 생성물과 CO2로 분해(3탄소 생성물은 엽육세포로 되돌아 가고 CO2 는 Rubisco와 결합하여 갤빈회로로 들어감) C3 식물에 비해 많은 ATP가 요구되나 밝은 조건하에 사는 식물들은 충분한 ATP를 생성할 수 있다.
3.호흡(RESPIRATION)
세포의 연료 분자들 중 포도당의 생성 반응과 분해 반응을 비교해 보자.
생성 : (광합성) 6CO2 + 6H2O + 에너지 → C6H12O6 + 6O2
△G = +686 kcal / mole
분해 : (호흡) C6H12O6 + 6O2 → 6CO2 + 6H20 + 에너지
△G = -686 kcal / mole
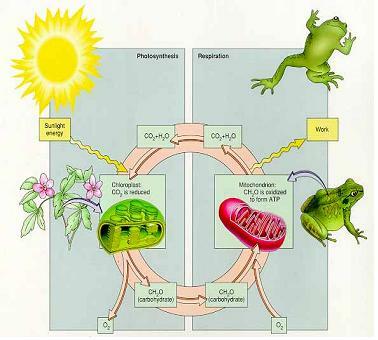
생명 진화 초기에 세포막 내의 수소이온(양성자) 농도가 높아짐, 수소이온을 퍼내는 역할을 ATP합성효소가 맡음. 세포막내에 많아진 수소이온을 pumping하는데 ATP를 소모.
ATP는 다른 생명현상에도 꼭 필요하기에 ATP를 아끼고 양성자를 pumping하는 다른 메카니즘을 만들어냄. 이온채널들이 양성자 pump역할을 할 수 있게 함 ⇒ 피브르산과 지방산이 해당과정을 통해 Acetyl CoA가 되고 TCA cycle를 돌아서 그 부산물인 NADH를 생성하여 NAD+H++2e- 가 될 때 high energy electron이 나온다. 전자의 고에너지를 이용해서 양성자를 pumping. 에너지를 잃은 e-를 담는 그릇이 O2이다.
O2 + 4e- + 4H+ → 2H2O
호흡이 왜 중요한가? 궁극적으로 양성자 pump를 사용하기 위해 high energy electron을 써야하고 에너지를 잃은 electron을 회수하기위해 O2가 필요하고 부산물로 나온 것이 H2O. 광합성과 호흡의 주역은 ATP합성효소이다.
1) THREE WAYS OF UTILIZING GLUCOSE
산소호흡aerobic respiration, 무산소호흡anaerobic respiration, 발효fermentation
(1) 해당과정glycolysis : 포도당 대사의 첫번째 단계. 기질수준인산화에 의해 ATP 생성
(2) 산소호흡과 무산소호흡
산소호흡 : 해당과정의 산물들이 산화되고 전자와 양성자는 전자전달계로 이동
→ 전자전달계를 통과한 전자는 산소와 결합(산소가 최종 전자 수용체)
화학삼투적 인산화에 의해 대부분의 ATP가 생성
무산소호흡 : 박테리아 종류(산소에 의해 해를 입는다) 최종 전자 수용체는 황산염 또는 질산염, 이산화탄소 최종 산물 - 아질산염, 아산화질소, 황화수소, 메탄가스
ATP의 대부분은 화학삼투적 인산화에 의해 생성
(3) 발효fermentation : 해당과정의 최종산물이 호흡으로 들어가지 않고 에너지가 풍부한 유기노폐물로 전환 → 전자전달계와 화학삼투적 인산화 없이 ATP 생산
(4) 호흡의 3 단계
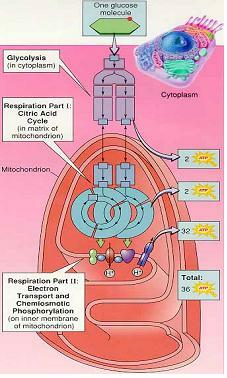
1 단계 : 해당과정(세포질)
- 포도당의 분해와 기질수준 인산화
2 단계 : 시트르산회로(미토콘드리아)
- 전자와 양성자를 생산하면서 연료가 CO2와 H2O로 완전히 산화
3 단계 : 전자전달과 화학삼투적 인산화 - 전자의 자유에너지를 이용해 양성자 기울기를 만들고 ATP 생성
2) GLYCOLYSIS
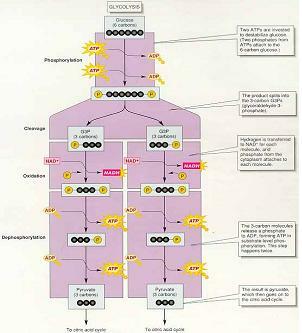
세포질에서 일어나는 호흡의 예비단계로 산소를 필요로하지 않는다.
포도당에서 시작하여 최종산물로 두 개의 피루브산을 만든다.
(조효소인 NAD+ 2분자가 NADH로 환원되고, 4분자의 ADP가 인산화되어 4ATP가 생성
→ 포도당 자유에너지의 일부는 NADH와 ATP에 보존)
(1) 해당과정의 핵심 반응
(2) 해당과정의 조절
세포가 빠른 속도로 ATP를 사용하면 해당과정도 빨리 진행
과당인산키나아제phosphofructokinase : 알로스테릭 효소로 해당과정의 반응 3.에서 과당-6-인산fructose-6-phosphate을 과당-1,6-인산fructose-1,6-diphosphate 로 전환시킨다.
세포내의 ATP 농도가 높으면 억제되고, ADP나 AMP 농도가 높으면 활성화
시트르산에 의해서도 억제 : 시트르산의 생성은 해당과정의 피루브산의 양에 달려있음
⇒ 시트르산이 과당인산키나아제를 억제 → 피루브산의 농도가 떨어짐
3) AEROBIC REPIRATION : THE MITOCHONDRION
(1)피루브산에서 아세틸-CoA로, NADH는 미토콘드리아로 들어갈 수 없지만 왕복운반자shuttle carrier에 의해 에너지를 가진 전자를 미토콘드리아로 전달, 미토콘드리아로 들어간 피루브산은 거대효소/조효소 복합체(티아민, 리보플라빈, 판토텐산, 니코틴산)에 의해 에너지가 풍부한 아세틸-CoAacetylcoenzyme A라는 기질의 일부분이 된다.
피루브산 + NAD+ + CoA → 아세틸-CoA + NADH + H+ + CO2
(2) 시트르산 회로 Citric acid cycle : H.A. Krebs
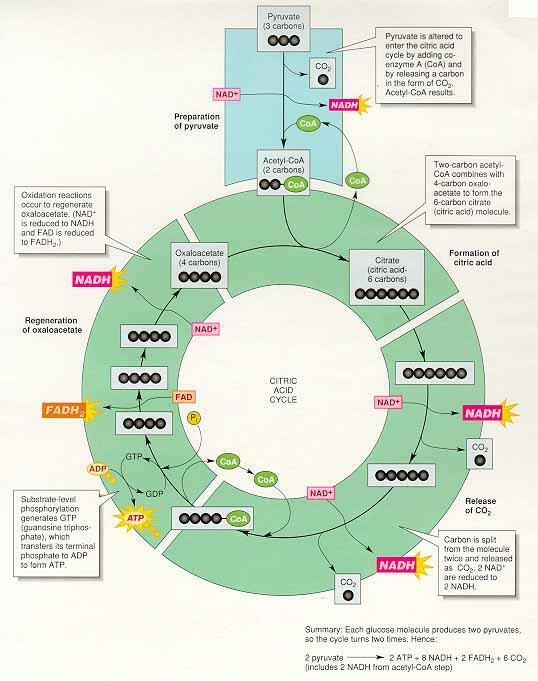
산화반응은 NAD+와 FAD와 같은 조효소의 환원과 함께 일어남
조효소는 전자와 양성자를 전자전달계로 이동 → 양성자 기울기 형성 → 화학삼투적 인산화에 작용
① 시트르산 회로의 주요 반응
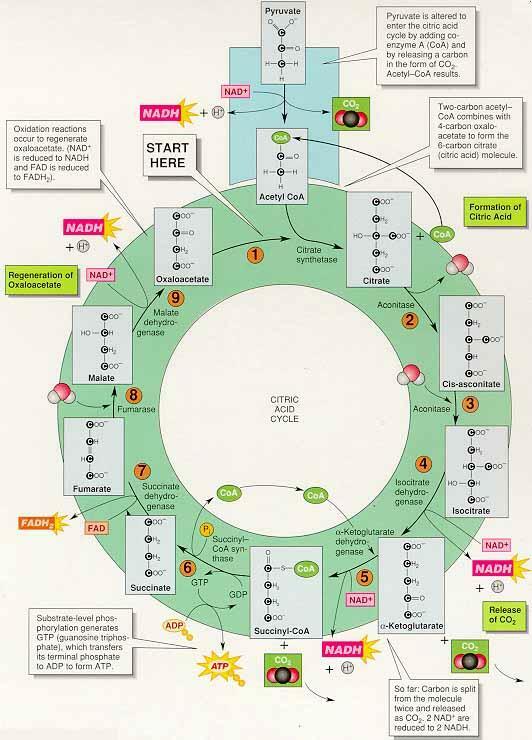
아세틸-CoA의 도입 : 옥살아세트산oxaloacetate과 반응하여 시트르산 생성, 시트르산 합성효소citrate synthesis가 관여하여 시트르산 회로로 들어감, 떨어져 나온 CoA는 다시 피루브산과 결합하여 아세틸-CoA를 생성하도록 재순환
이산화탄소의 방출 : 순환하기 위해 마지막 단계의 옥살아세트산이 형성되어야 하는데 6탄소를 갖는 시트르산이 4탄소의 옥살아세트산으로 전환되기 위해 서 두 번의 탈카르복시화(이산화탄소 제거)가 일어남
시트르산 회로에서의 산화 : 연료의 산화는 조효소 NAD+와 FAD가 전자와 양성자를 받아 환원되는 반응과 짝을 이룸, 환원된 조효소는 전자전달계와 양성자 펌프를 활성화시킴
기질수준의 인산화 : 반응 6.에서 해당과정과 마찬가지로 기질수준 인산화substrate-level phosphorylation가 일어남, GDPguanosine diphosphate는 인산화되어 GTPguanosine triphosphate가 생성 → GTP는 다시 ADP를 인산화 하여 ATP를 생성, GTP는 화학적으로 ATP와 유사하지만 질소염기가 아데닌이 아니라 구아닌이다.
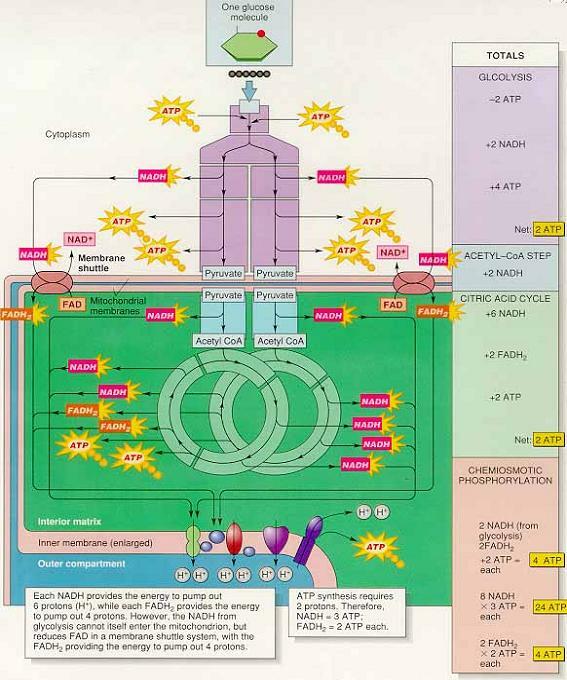
② 지금까지의 포도당 이화작용 : 호흡은 포도당에서 시작되고 포도당은 두 분자의 피루브산 생산 ⇒ 2ATP, 2NADH, 6NADH, 2FADH2 생성
4) Electron Transport Systems and Chemiosmotic phosphorylation
(1) 미토콘드리아의 구조
외막-외부와 내부로 물질을 능동수송하거나 왕복시키는 수송단백질transport protein이 존재
외막과 내막 사이에 외부구획outer compartment이 존재
내막-내부구획inner compartment인 기질matrix을 둘러싸고 있음
크리스테cristae 형성 - 내막의 표면적을 증가시켜 반응면적을 증가
① 전자전달계와 F0 F1 복합체
내막에 많은 전자전달계가 있어서 전자를 전달
양성자 펌프(양성자를 내부구획에서 외부구획으로 수송)의 운반자는 철을 가진 시토크롬
F0 복합체 : 외부구획에서 시작해 내부구획으로 돌출
F1 복합체 : ATP 합성효소를 포함하고 있어 화학삼투적 인산화에 중요
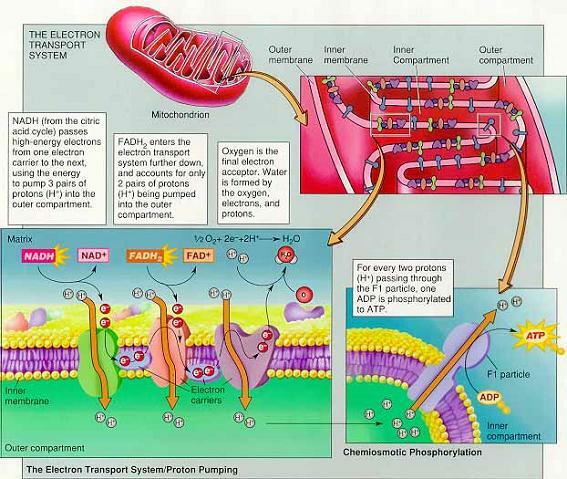
(2) NADH의 전자가 화학삼투 기울기 형성
시트르산 회로에서 생성된 NADH와 FADH2의 화학삼투 기울기 형성 ⇒ NADH가 전자전달계의 첫번째 운반자 FMN에 도착하면서 시작, FADH2는 CoQ에 작용하여 두번째 단계의 양성자 펌프에서부터 시작.
NADH가 두 개의 전자와 양성자를 막횡단 단백질인 FMN(flavin monomucleotide)에 넘겨주는 순간 전자전달계에서 전자의 흐름 시작(FMN이 첫번째 양성자 펌프)
두 전자가 CoQ(조효소 우비쿼논)로 전달 → CoQ는 내부구획의 두 양성자를 잡아 CoQH2가 되고 외부구획으로 방출하면서 시토크롬b(cyt b)로 전자를 전달 (FADH2의 작용은 CoQ에서부터 양성자 펌프 시작) 시토크롬b는 두 개의 양성자를 외부구획으로 수송하며 두 전자를 시토크롬c(막 표면에 위치)로 전달 → 시토크롬a에 의해 전자는 기질의 산소로 전달되어 물 형성 (산소는 마지막 전자수용체)
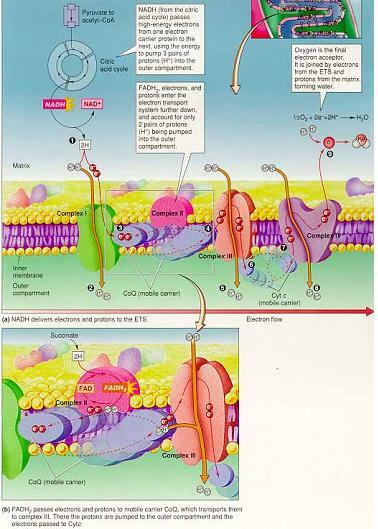
① 산소의 역할
호흡의 최종단계에서 자유에너지의 대부분을 소모한 전자와 결합.
한 분자의 산소를 환원시키기 위해서는 두 개 조효소로부터 전자가 와야함
O2 + 4e- + 4H+ → 2H2O
NADH에서 시작해 산소에까지 이르는 반응 → 발열반응(-53kcal/mole)
⇒초기 상태의 자유에너지는 양성자 기울기의 자유에너지로 축적
② 양성자 생성 개요
미토콘드리아로 들어간 두 분자의 피루브산은 8개의 NADH와 2개의 FADH2를 생성
NADH의 전자는 6개의 양성자를 펌프하고, FADH2의 전자는 4개의 양성자를 펌프 ⇒ 두 분자의 피루브산은 56개의 양성자를 외부구획으로 펌프(8×6 + 2×4)
해당과정에서 생성된 세포질에 있는 2개의 NADH는 미토콘드리아 외막에 있는 운반자에 의해 미토콘드리아 조효소(FAD, NAD+)를 환원시켜 2개의 FADH2를 생성 ⇒ 8개의 양성자 펌프 ∴ 64개의 양성자가 펌프됨
(3) 화학삼투 기울기와 ATP 생성의 개요
두 개의 양성자가 F1복합체를 통과할때 한 분자의 ATP 생성 (포도당이 완전히 산화되면 36개의 ATP 생성)
포도당 한 분자당 생기는 총 ATP 수
- 기질수준 인산화 : 해당과정 2ATP, 시트르산회로 2ATP
- 화학삼투 인산화 : 64 양성자 → 32ATP
- 포도당 한 분자당 총계 : 36 ATP
(4) 양성자 기울기의 다른 사용
인산화에 필요한 인산(Pi)을 미토콘드리아로 이동시키는 데 이용, 미토콘드리아로 칼슘을 이동시키는데 사용되기도 함, 인산과 칼슘은 특별한 공동왕복 운반자에 의해 수송되며 양성자와 함께 이동, 양성자 기울기는 ADP, 인산, 칼슘 수송에 일상적으로 이용
5) Fermentation Pathways
전자전달계와 화학삼투적 인산화가 관여하지 않음
(1) 알콜 발효alcoholic fermentation - 효모와 몇몇 박테리아에서 발생
최종산물 : NAD+, 이산화탄소, 에틸알코올
피루브산이 탈카르복시화 효소의 작용으로 아세트알데히드 생성, CO2방출
→ NADH가 알코올 탈수소효소에 의해 산화
→ 전자와 양성자가 아세트알데히드로 전달되어 에틸알코올을 형성
※ 효모 : 조건 무산소성 생물 facultative anaerobes - 조건에 따라 무산소성 알코올 발효 또는 이산화탄소와 물이 최종 산물인 산소호흡 중 하나를 택해 피루브산 대사
(2) 젖산 발효lactate fementation - 박테리아와 동물 근육세포의 중요한 경로
최종산물 : NAD+와 젖산
① 근육조직의 해당과정과 젖산 발효
심한 운동으로 인한 많은 양의 ATP 소모와 산소 공급의 부족
(근육세포) ⇒ 1. 근육의 풍부한 크레아틴인산creatin phosphate에 고에너지 인산을 저장.
크레아틴인산은 근육 수축에 필요한 에너지를 직접 제공하지는 않지만 자신의 고에너지 인산기를 ADP에 전달하여 ATP 생성 : 크레아틴인산 + ADP → ATP + 크레아틴
2. 크레아틴인산이 점차 소모되면 사용되는 두번째 방법인 해당과정 시작. 그러나 산소가
공급되지 못하면 NAD+의 재생이 일어나지 못해 해당과정이 진행되지 못함
⇒ 임시 방법으로 NAD+를 재생산
NADH에서 수소를 받아 피루브산은 젖산으로 환원 → 젖산은 근육에 의 해 직접 사용될 수
없기 때문에 축적(일부는 간으로 보내짐) → 산소가 충분히 공급되면 젖산은 다시 피루브산이되고 산소호흡 경로로 들어감
※ 간으로 보내진 젖산은 피루브산으로 다시 전환된 후 포도당 신생합성 gluconeogenesis 경로로 들어간다 ⇒ 젖산은 포도당으로 전환되고, 포도당의 일부는 다시 근육으로.
② 산소부채oxygen debt : 산소와 크레아틴인산이 부족한 상태
운동하기 전의 상태로 회복되는데 필요한 산소의 양으로 표현
※포도당 대사의 요약
6) Alternative Fuels for the Call
지방과 단백질, 탄수화물은 중간대사intermediary metobolism 경로에 의해 해당과정과 시트르산 회로에 사용할 수 있도록 소단위로 분해
(1) 지방 대사
지방산이 미토콘드리아 막을 지나 내부구획으로 수송되기 위해서는 조효소A(CoA)에 의해 활성화 ⇒ 지방산은 CoA와 결합하여 기질로 운반
→ 아세틸-CoA(2C)가 떨어져 나와 시트르산 회로로 들어감
→ 다시 지방산과 CoA가 다시 결합하고 아세틸-CoA가 떨어져 나오는 과정 반복하여
지방산이 완전히 분해
(2) 단백질의 준비
소화효소에 의해 20개의 아미노산으로 가수분해
→ 탈아미노화반응deamination에 의해 아미노기 제거, 5개는 피루브산으로 전환 후 다시 아세틸-CoA로 전환, 6개는 피루브산 단계를 건너 뛰어 아세틸-CoA로 시트르산회로에 들어감, 나머지 아미노산은 시트르산회로를 구성하는 특정한 산으로 전환되어 시트르산회로로 들어감
(3) 중간대사 - 생합성 경로
생합성biosynthesis 경로 : 시트르산 회로의 호흡에 대한 중심적 역할 외에 다른 반응에 필요한 원료물질을
제공하는 중간대사의 한 부분
4. 리처드 파인만
(중력과 원자력을 제외하고) 우주에서 일어나는 모든 현상은 3가지로 설명가능
하나. 광자가 여기에서 저기로 움직인다.
둘. 전자가 여기에서 저기로 움직인다.
셋. 전자가 광자를 흡수하거나 방출한다.
대표적 예가 광합성이나 호흡에서의 전자전달계.
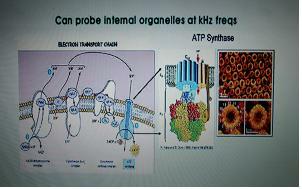
⇒ 생명의 출발점은 electron가 photon을 흡수하면서 시작. 빛에너지를 전자가 받아서 그 에너지를 단백질 전달 시스템을 통해 전달되고 그 과정 중 수소이온이 흡입, 다시 수소이온을 pumping하면서 ATP생성
5.ATP 합성효소 - 거대한 나노머신(35억년)
ATP [adenosine triphosphate]
아데노신에 인산기가 3개 달린 유기화합물로 아데노신3인산이라고도 한다. 이는 모든 생물의 세포 내 존재하여 에너지대사에 매우 중요한 역할을 한다. 즉, ATP 한 분자가 가수분해를 통해 다량의 에너지를 방출하며 이는 생물활동에 사용된다.

양성자가 들어오면서 축이 돌아가고 ADP가 Pi와 합해져서 ATP를 합성(터빈과 같다)
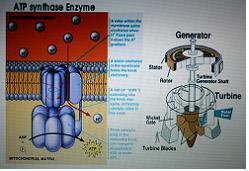
생명현상 초기에 나온 머신이며 지금도 이보다 복잡한 나노머신은 없다.