 제18강 운 동 출 력
제18강 운 동 출 력
-뇌의 모든 계산결과, 감각입력, 운동출력이 최종적으로 표출되는 것은 우리 몸을 구성하고 있는 골격근이다.
-운동출력이 마음의 출현과 어떤 관련이 있을까? 마음의 출현을 이끄는 전체과정에서 감각입력보다도 운동출력을 중심으로 이해하는 게 더 효율적이다. 근육운동을 통해 마음의 형성을 추적해나갈 때 문제의 본질에 가까워진다.(꿈꾸는 기계의 진화-단세포의 미오닉 운동을 통해 감정이나 언어까지도 유추해 볼 수 있다고 주장)
1.골격근
네 가지 주요 조직유형중 하나인 근조직은 주로 근세포(통상 근섬유(muscle fibers)라 한다)로 구성되어 있으며, 이 세포들은 수축을 위해 고도로 분화되어 있다. 근조직에는 골격근(skeletal muscle), 심근(cardiac muscle), 평활근(smooth muscle)의 세 가지 유형이 있다. 골격근조직은 결합조직, 신경 및 혈관들도 함께 포함되어 있는 기관인 골격근(skeletal muscle)에서 발견된다. 골격근이라는 이름이 암시하듯이, 골격근은 직접 또는 간접적으로 골격의 뼈들에 부착하고 있다. 골격근조직은 골격근에 다음과 같은 기능을 수행할 수 있는 능력을 부여한다.
1.골격 움직임을 일으킨다(Produce skeletal movement). 골격근의 수축은 건을 당겨 골격의 뼈를 움직인다. 이 작용의 범위는 팔을 뻗는 것과 같은 단순한 동작부터 수영, 스키, 타이핑과 같이 고도로 협응된 움직임에 이르기까지 매우 넓다.
2. 자세와 신체의 위치를 유지시킨다(Maintain posture and body position). 골격 근내 장력이 독서시 머리가 한자리에 고정되게 하고 보행시 양 발이 체중을 싣고 균형 잡을 수 있게 하는 등 신체의 자세를 유지시킨다. 지속적인 근육의 활동이 없다면 우리는 무너짖 않고 똑바로 앉을 수 없으며, 비틀거리지 않고 서있을 수도 없다.
3. 연부조직을 지지한다(Support soft tissue). 복벽과 골반강의 바닥은 골격근층으로 이루어져 있다. 이 근육들은 여러 내장의 무게를 지탱하고 내부조직을 상해로부터 보호해준다.
4. 출입을 통제한다(Guard entrances and exits). 소화관과 요로의 출입구는 골격근에 의해 둘러싸여 있다. 이 근육들은 연하, 배변, 배뇨가 수의적으로 제어되게 한다.
5. 체온을 유지한다(Maintain body temperature). 근수축은 에너지를 필요로 하고, 체내에서 에너지가 사용되는 곳에서는 모두 약간의 에너지가 열로 전환된다. 활동중인 근육에서 유리된 열이 정상 기능활동에 필요한 범위내에서 체온을 유지시킨다.
■ 골격근 해부학(Anatomy of Skeletal Muscle)
그림 10-1은 대표적인 골격근의 모양과 체제를 도해하고 있다. 골격근은 결합조직, 혈관, 신경과 함께 골격근조직을 내포하고 있다.
□ 결합조직 체제(Connective Tissue Organization)
세 가지 결합조직층-바깥의 근외막(epimysium), 가운데 근다발막(perimysium), 안의 근내막(endomysium)-은 모든 근육의 한 부분이다. 이 층들과 그들의 관계가 그림 10-1에 도해되어 있다. 근육 전체는 치밀교원섬유층인 근외막(epimysium)으로 둘러싸여 있다. 근외막은 근육을 주위의 조직들 및 기관들과 격리시킨다.
근다발막(perimysium)의 결합조직섬유들은 골격근을 일련의 구획들(compartments)로 나누며, 각각의 구획들은 근다발(fascicle)이라 하는 근섬유 다발을 수용하고 있다. 근다발막은 교원섬유와 탄성섬유와 함께 근다발에 혈류를 유지시키고 신경지배하는 혈관과 신경들을 수용하고 있다.
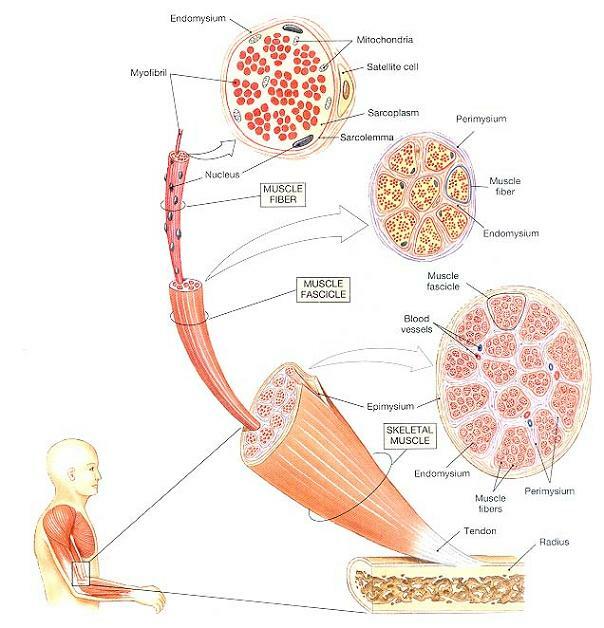
그림 10-1. 골격근의 체제(Organization of Skeletal Muscles).
골격근은 근외막으로 에워싸여져 있는 근다발들로 구성된다. 근다발들은 결합조직섬유로 된 근다발막에 의해 격리되며, 각각의 근다발내에는 근섬유들이 근내막에 의해 둘러싸여 있다. 각각의 근섬유는 많은 천층 핵들뿐만 아니라 그림 10-2에서 자세하게 나타내진 미토콘드리아 및 기타 세포소기관들도 갖고 있다.
근다발내에서는, 섬세한 결합조직인 근내막(endomysium)이 골격근섬유를 둘러싸며 인접 근섬유들과 묶어 놓는다. 흩어져 있는 성상세포(satellite cells)는 근내막과 근섬유 사이에 위치한다. 이 세포들은 손상된 근조직을 복구하는 기능을 한다.
근내막과 근다발막의 교원섬유들은 서로 얽혀 있으며, 근다발막의 교원섬유들은 근외막의 근외막의 섬유들과 서로 얽혀 있다. 근육의 양끝에서, 근외막의 교원섬유들은 서로 합쳐져 건(tendon)이라 하는 다발이나 건막(aponeurosis)이라 하는 넓은 시트(sheet)를 형성한다. 치밀정형결합조직의 예인 건과 건막은 골격근을 뼈에 부착시킨다. 건의 섬유들은 근외막과 서로 얽혀 있으며 골간질내로 뻗어나가 단단한 부착을 제공한다. 그 결과, 근육의 모든 수축은 건과 건이 부착되어 있는 뼈를 당기는 힘을 발휘한다.
□ 혈관과 신경(Blood Vessels and Nerves)
근외막과 근다발막의 결합조직은 근섬유들에 공급되는 혈관과 신경을 수용하고 있다. 근수축은 막대한 양의 에너지를 필요로 한다. 대규모의 혈관망이 산소와 양분을 공급하고 활동하는 골격근에 의해 산출된 대사폐기물들을 실어나간다. 대부분의 경우 혈관들은 신경과 함께 근육으로 들어가고, 혈관과 신경은 근다발막까지 동일한 패턴으로 가지를 친다. 일단 근내막안에 들어가면, 소동맥은 개개의 근섬유들을 둘러싸고 있는 모세혈관망에 혈액을 공급한다.
골격근은 오직 중추신경계(CNS)에서 오는 자극에 의해서만 수축한다. 근외막을 관통하는 축색(axon), 즉 신경섬유(nerve fibers)는 근다발막을 통해 가지를 치고 근내막으로 들어가 개개의 근섬유들을 지배한다. 골격근은 종종 수의근(voluntary muscles)이라 불리는데, 이들의 수축을 수의적으로 조절할 수 있기 때문이다. 아울러 이 골격근중 많은 수가 불수의적으로 조절되기도 한다. 예를 들면, 횡격막과 같이 호흡에 관여하는 골격근들은 보통 불수의적인 조절 하에 움직인다.
□ 골격근섬유의 현미경해부학(Microanatomy of Skeletal Muscle Fibers)
골격근섬유는 전형(typical)적인 세포와는 무척 다르다. 확실한 차이의 하나는 크기로, 골격근섬유들은 거대하다. 대퇴근의 근섬유는 직경이 100㎛이고 길이는 전체근육의 길이(30~40㎝)와 같다. 두 번째 명백한 차이점은 골격근 섬유가 다핵이라는 점이다. 즉, 각각의 골격근섬유들은 세포막 바로 밑에 수백개의 핵을 수용하고 있다. 이 핵들 속에 내포된 유전자들은 정상 수축에 필요한 효소와 구조단백질의 생산을 지시하며, 이 유전자들의 복사본들(copies)이 다수로 존재하여 그 과정의 속도를 높인다. 이것은 매우 빠르게 대사전환이 일어나는 골격근섬유에서 특히 중요하다.
독특하게 큰 모습과 다핵이란 특성은 서로 연계되어 있다. 발생기 동안 근원세포(myoblasts)라 하는 배아의 세포군이 서로 융합하여 그림 10-2에 나타내져 있는 것과 같은 개개의 골격근섬유들을 만들어낸다. 골격근섬유의 핵들은 각기 참여한 근원세포들을 반영한다. 어떤 근원세포들은 발생중인 근섬유들에 융합하지 않는다. 이 융합하지 않은 세포들은 그림 10-1에서 보이는 것처럼 성장된 골격근섬유에서 성상세포로 남는다. 손상을 받은 후, 성상세포들은 커지고 분열하여 손상된 근섬유들과 융합함으로써 조직의 재생을 돕는다.
-당신들을 이루는 세포들 자체가 우리 세균들이 10억년전에 발견한 낡은 기술들을 똑같이 재현하는 세균균체들이다. - 리처드 도킨스의 <조상 이야기>
-근세포는 아주 특이화된 세포. 단백질사슬을 한방향으로 나열, 움직임에 특화된 세포
근초와 T세관(Sarcolemma and Transverse Tubles)
근섬유의 세포막, 즉 근초(sarcolemma)는 세포질, 즉 근형질(sarcoplasm)을 둘러싸고 있다. 다른 세포막과 마찬가지로, 근초는 특징적인 막전위를 갖고 있다. 골격근섬유에서, 갑작스럽게 전도된 막전위의 변화가 수축을 이끌어내는 첫 번째 단계이다. 골격근섬유는 매우 크기 때문에, 수축을 위한 신호는 세포의 안쪽에도 전달되어야 한다. 이것은 횡세관(transverse tubule), 즉 T세관에 의해 일어난다. T세관은 근초에서 시작하여 막표면과 직각으로 근형질 안으로 뻗어 있는 좁은 관이다. T세관은 세포외액으로 채워져 있고, 이들은 산을 관통하는 터널들처럼 근섬유를 관통하는 통로를 형성한다. T세관은 근초에서 열려져있고, 각각의 세관을 이루는 막들은 근초와 전체적으로 동일한 성질을 갖고 있다.
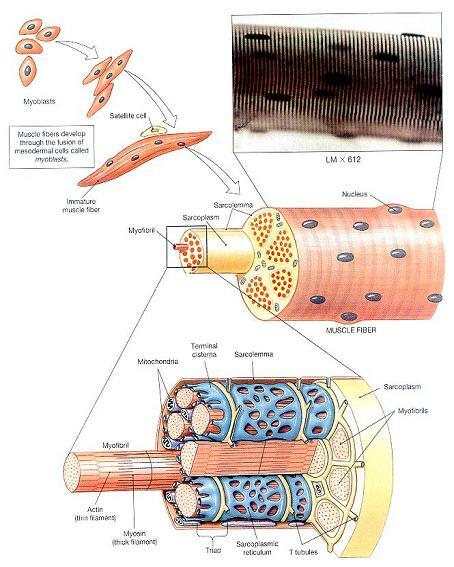
그림 10-2. 골격근섬유의 형성과 구조(Formation and Structure of a Skeletal Muscle Fiber).
근원섬유(myofibrils)
근섬유내에서, T세관의 가지들은 근원섬유(myofibrils)라 하는 원통형 구조물을 에워싼다. 근원섬유(그림 10-2)는 직경 1~2㎛ 정도이고 길이는 세포 전체의 길이와 같은 원통형 구조이다. 각각의 골격근섬유는 수백~수천 개의 근원섬유를 수용하고 있다.
근원섬유는 주로 액틴(actin)과 미오신(myosin)으로 구성된 단백세사인 근세사(myo- filaments)다발이다. 근원섬유는 능동적으로 짧아질 수 있다. 즉, 이들은 골격근섬유의 수축을 담당하는 세포소기관(organelles)이다. 이 단백질들은 세포막 내면과 결합될 수 있다. 골격근섬유의 양 끝에서, 근세사는 근초에 부착된다. 그 결과, 근세섬유가 수축하면 세포 전체가 짧아진다. 근원섬유 사이에는 미토콘드리아와 포도당의 저장형인 글리코겐 과립들이 흩어져 있다. 포도당은 해당(glycolysis)을 통해 분해되고 미토콘드리아의 활동은 활기찬 근수축에 필요한 ATP를 제공한다.
근형질세망(The Sarcoplasmic Reticulum)
T세관이 근세섬유를 둘러싸는 모든 곳에서, T세관은 근형질세망(sarcoplasmic reticu- lum)의 막과 밀착한다. 근형질세망(SR)은 다른 세포의 활면내형질세망(SER)과 유사한 막 복합체이다. 골격근섬유에서, 근형질세망은 개개의 근원섬유를 덮어씌우는 세관망(tubular network)을 형성한다.(그림 10-2와 10-3). T세관의 양 옆에서, 근형질세망의 관들은 커지고 융합되어 종말소조(terminal cisternae)라 하는 확대된 방을 형성한다.
한 쌍의 종말소조에 T세관을 더한 조합을 트라이어드(triad)라 한다. 비록 이 트라이어드의 막들은 밀착하고 있어 서로 단단히 결합되어 있지만, 이들 사이의 직접적인 연결은 없고, 이들의 액체 내용물들도 분리되어 별개로 남아있다.
대부분의 세포들은 칼슘이온을 세포막을 통해 세포외액으로 펌프질해낸다. 비록 골격근섬유들도 이런 방식으로 세포 바깥으로 칼슘이온들을 펌프질해 내지만, 아울러 이들은 칼슘이온을 근형질세망(SR)의 소조(cisternae)로도 능동수송하여 세포질에서 칼슘이온을 제거한다. 휴지기의 골격근섬유의 근형질은 약 10-7m㏖/l정도의 매우 낮은 농도로 칼슘이온을 수용한다. 종말소조(terminal cisternae) 내의 유리 칼슘이온 농도는 이보다 약 1000배정도 더 높다. 아울러 소조는 칼슘이온과 가역적으로 결합하는 단백질인 칼세퀘스트린(calsequestrin)이라는 단백질을 수용하고 있다. 유리칼슘과 결합칼슘을 고려한다면, 소조내의 총 칼슘이온 농도는 주위의 근형질내의 농도보다 40,000배정도 더 많을 수도 있다.
저장된 칼슘이온이 근형질내로 유리될 때 근수축이 시작된다. 그러면 칼슘이온은 근절(sarcomeres)이라 하는 개개의 수축단위내로 확산된다.
근절(Sarcomeres)
근원섬유는 액틴과 미오신 근세사로 된 다발이다. 액틴은 가는 세사(thin filaments)에서 발견되고, 미오신은 두터운 세사(thick filaments)를 형성한다. 두 근세사는 근절(sarcomeres)이라 하는 반복되는 기능단위로 조직화되어 있으며, 이는 그림 10-3에 상세히 나타내져 있다.
근절의 체제(Sarcomere Organization)
근원섬유는 대략 10,000개의 일직선으로 늘어선 근절들로 구성되어 있으며, 각각의 근절들은 1.6-2.6μm의 휴지기 길이를 가지고 있다. 근절은 근섬유의 가장 작은 기능단위로, 근절의 두터운 세사와 가는 세사의 상호작용이 근수축을 담당한다. 근절은 ⑴두터운 세사, ⑵가는 세사, ⑶두터운 세사와 가는 세사의 위치를 안정시키는 단백질 및 ⑷두터운 세사와 가는 세사 사이의 상호작용을 조절하는 단백질을 내포하고 있다.
그림 10-3은 개개의 근절의 구조를 보여주고 있다. 두터운 세사와 가는 세사의 크기, 밀도 및 분포의 차이가 근절의 띠모양이 나타나게 한다. 어두운 부분, 즉 두터운 세사 전부와 가는 세사 일부를 수용하는 부분을 A대(A-band)라 한다. 가는 세사만을 수용하는 밝은 부분은 I대(I-band)라 한다. (A-band와 I-band라는 용어는 anisotropic(異方性), isotropic(等方性)에서 유래되었는데, 이는 편광하에서 볼 때 나타나는 이 대들의 형태를 언급한 것이다.) A대는 dark의 A를 생각해 어둡다, I대는 light의 I를 생각해 밝다고 하면 기억하는데 도움이 될 것이다.

그림 10-3. 근절의 구조(Sarcomere Structure).
⒜ 근절(sarcomere)과 트라이어드(triad) 사이의 관계.
⒝ 장딴지에 있는 비복근의 근섬유에서 취한 근원섬유의 근절. (TEM × 64,000)
⒞ 근절내 가는 세사와 두터운 세사의 체제.
⒟ 근절내 여러 부위의 단면.
▪A대(A Band). 두터운 세사는 A대 안에서 근절의 중앙에 위치한다. A대의 길이는 전형적인 두터운 세사의 길이와 동일하다. A대는 다음 몇 가지로 세분되어진다.
1.M선(M line). 각각의 두터운 세사의 중앙부는 M선의 단백질에 의해 양쪽이 연결된다. 이 짙게 염색되는 단백질들은 두터운 세사의 위치를 안정시키는데 도움을 준다.
2.H구역(H zone). 휴지기의 근절에서 H구역, 즉 H대(H band)는 M선 양쪽의 밝은 부분이다. H구역은 두터운 세사만을 수용하고 가는 세사는 수용하지 않는다.
3.겹친 구역(Zone of overlap). 겹친 구역에서는 두터운 세사 사이에 가는 세사가 확인된다. 이 지역에서 각각의 가는 세사는 세 개의 두터운 세사에 의해 형성된 삼 각형 속에 위치하고, 각각의 두터운 세사는 여섯 개의 가는 세사에 의해 둘러싸인다.
▪I대(I Band). I대는 한 근절의 A대부터 다음 근절의 A대까지 펼쳐져 있다. Z선(Z line)는 인접한 근절 사이의 경계선을 표시한다. Z선은 커넥틴(connectins)이라 하는 단백질로 구성되어 있는데, 이것은 인접한 근절들과 연계된 가는 세사들을 서로 연결 시킨다. 가는 세사는 근절의 양끝 Z선부터 M선을 향해 뻗어 겹친 구역내로 들어간다.
각각의 근절마다 두 개의 T세관이 둘러싸고 있고, M선 양쪽 겹친 구역 부위에 트라이어드(triads)가 있다. 그 결과, 근형질세망(SR)에서 유리된 칼슘이온은 두터운 세사와 가는 세사가 상호작용할 수 있는 부위로 들어가게 된다.
근섬유 전체는 인접한 근원섬유의 Z선들이 정렬되어 있기 때문에 띠를 두른 모습으로 나타난다. 아울러 골격근조직을 횡문근(striated muscle)이라고도 하는데, 광학현미경으로 볼 때 이 띠들, 즉 횡문(striations)이 보이기 때문이다.
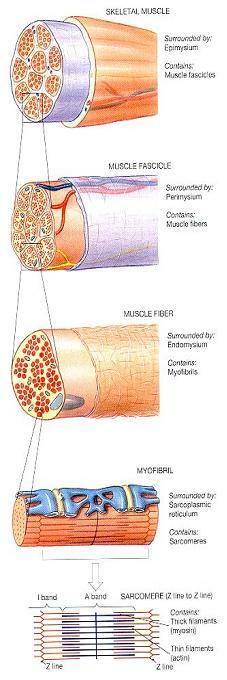
그림 10-4. 골격근섬유의 기능적 체제 수준(Levels of Functional Organization in a Skeletal Muscle Fiber).
그림 10-4는 이제까지 우리가 논의해왔던 체제 수준들을 다시 보여주고 있다. 이제 이 체제의 분자 수준을 조사하면서 근수축을 담당하는 근세사의 구조를 살펴보기로 하자.
가는 세사(Thin Filaments)
전형적인 가는 세사는 직경이 5-6㎚이고, 길이가 1㎛이다. 하나의 가는 세사는 세 가지 단백질-F 액틴(F actin), 트로포미오신(tropomyosin), 트로포닌(troponin)-을 내포하고 있다.
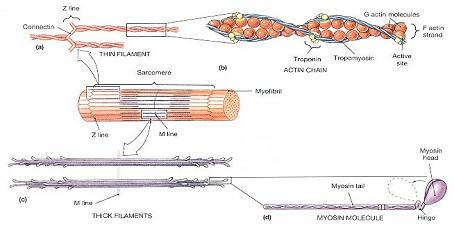
그림 10-5. 두터운 세사와 가는 세사(Thick and Thin Filaments).
⒜ 가는 세사의 거시 구조, Z선에의 부착을 보여주고 있다.
⒝ F 액틴 가닥내의 G 액틴 아단위들의 체제와 트로포닌-트로포미오신 복합체의 위치.
⒞ 두터운 세사의 구조, 두터운 세사를 따라 미오신 분자들의 방향을 보여주고 있다.
⒟ 미오신 분자의 구조
 ▪F액틴(F actin)은 구형 분자인 G액틴(G actin) 300~400개로 구성된 비꼬인 가닥이다. 각각의 G 액틴 분자들은 두터운 세사와 결합할 수 있는 활성 부위(active site)를 수용하고 있는데, 이는 기질 분자가 효소의 활성부위에 결합하는 것과 같은 방식이다. 휴지 상태에서는 트로포닌-트로포미오신 복합체(troponin-tropomyosin complex)에 의해 미오신 결합(myosin binding)이 방지된다.
▪F액틴(F actin)은 구형 분자인 G액틴(G actin) 300~400개로 구성된 비꼬인 가닥이다. 각각의 G 액틴 분자들은 두터운 세사와 결합할 수 있는 활성 부위(active site)를 수용하고 있는데, 이는 기질 분자가 효소의 활성부위에 결합하는 것과 같은 방식이다. 휴지 상태에서는 트로포닌-트로포미오신 복합체(troponin-tropomyosin complex)에 의해 미오신 결합(myosin binding)이 방지된다.
▪트로포미오신(tropomyosin) 가닥은 활성부위를 덮어 액틴-미오신 상호작용을 방지한다. (그림 10-5b) 트로포미오신 한 분자는 7개의 활성부위를 덮고 있는 하나의 단백질 가닥이다. 이것은 전체 길이의 중간에서 트로포닌 한 분자와 결합하고 있다.
▪트로포닌(troponin) 한 분자는 3개의 구형의 아단위로 구성되어 있다.
1.첫 번째 아단위는 트로포미오신과 결합해 이들을 서로 묶어놓는다.
2.두 번째 아단위는 G 액틴과 결합해 트로포닌-트로포미오신 복합체를 제 위치에 고정시킨다.
3.세 번째 아단위는 칼슘이온과 결합하는 수용체를 갖고 있다. 휴지상태의 근육에서는, 세포내 칼슘이온농도는 매우 낮아 결합부위는 비어 있다.
트로포닌-트로포미오신 복합체의 위치가 변하여 F 액틴
의 활성부위가 노출되지 않는 한 수축은 일어나지 않는다. 칼슘이온이 트로포닌 분자의 수용체에 결합할 때 수축에 필요한 위치 변화가 일어난다.
근절의 양쪽 끝에서, 가는 세사는 Z선에 부착되어 있다. (그림 10-5a) Z선은 근원섬유의 표면에서 짙은 선으로 보이기 때문에 선(line)이라고 불리우지만, 단면 모습(sectional view)에서의 Z선은 펼쳐진 그물(open meshwork)처럼 보인다. (그림 10-3d). 이런 까닭에 Z선은 종종 Z판(Z disc)이라고도 한다.
두터운 세사(Thick Filaments)
두터운 세사(그림 10-5c)는 직경이 10~12㎚이고, 길이는 1.6㎛이다. 각각의 두터운 세사는 대략 500개의 미오신 분자들로 구성되어 있다. 하나의 미오신 분자는 서로를 휘감고 있는 한 쌍의 미오신 아단위로 구성되어 있다.(그림 10-5d) 길고 부착되어 있는 꼬리(tail)는 두터운 세사 안의 다른 미오신 분자와 결합하고 있다. 부착되지 않은 구형의 미오신 머리(head)는 가장 가까운 가는 세사를 향해 밖으로 돌출되어 있다. 수축시 미오신 머리는 가는 세사와 상호작용하기 때문에, 이들을 연결다리(cross-bridges)라고도 한다. 머리와 꼬리 사이의 연결은 경첩같이 기능하여 머리가 기저부에서 선회할(pivot) 수 있게 해준다. 선회(pivoting)가 일어나면, 머리는 M선을 향하거나 멀어지게 호를 그리며 움직인다(swing). 이 선회가 근수축의 핵심 단계이다.
모든 미오신 분자들은 꼬리가 근절의 중앙을 향하도록 배열되어있다.(그림 10-5c) 휴지상태의 근절의 H구역은 미오신 머리가 없는 부위에 해당한다. H구역 밖에서 미오신 분자들은 나선형으로 배열되어 있는데, 미오신 머리들은 주위의 가는 세사의 G 액틴 분자들을 향하고 있다. (그림 10-3d, 10-5c).
세사활주 이론(The Sliding Filament Theory)
골격근섬유의 수축을 지켜보는 동안 몇 가지 중요한 주목할 만한 사항들이 관찰된다. 수축이 일어날 때, H구역과 I대는 작아지고, 겹친 구역(zones of overlap)은 커지며, Z선은 서로 가까워지게 움직인다. A대의 넓이는 수축하는 동안 일정하게 유지된다. 오직 가는 세사가 근절의 중심을 향해 두터운 세사와 평행선을 유지하며 미끄러져 들어가는 경우만 이 관찰결과와 들어맞는다. 이 활주 작용은 그림 10-6에 도해되어 있다. 근수축 과정 동안 일어나는 생리적 변화에 대한 이 설명을 세사활주 이론이라 한다.
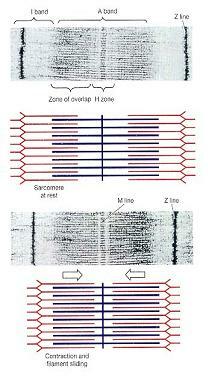
그림 10-6. 골격근 수축시 근절의 형태 변화(Changes in the Appearance of a Sarcomere). 수축시 A대는 동일한 넓이를 유지하지만, Z선은 서로 가까워지고 I대는 작아진다.
2.골격근의 수축(Contraction of Skeletal Muscle)
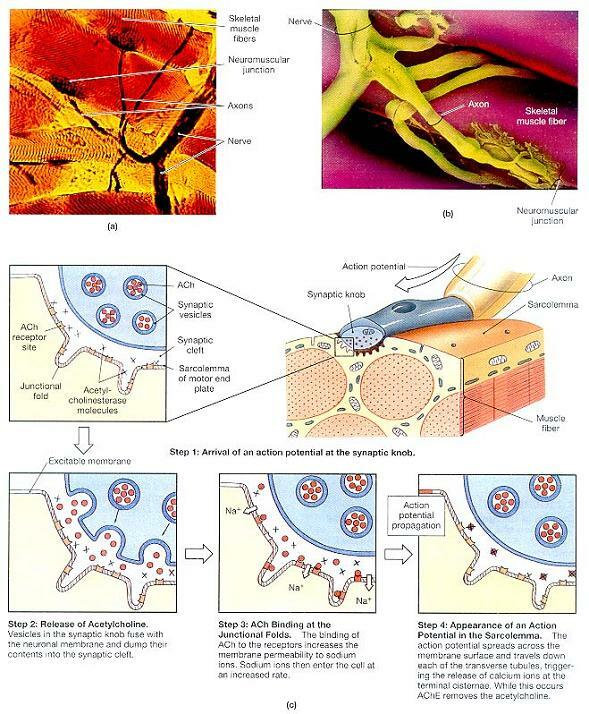
그림 10-7. 골격근 신경지배(Skeletal Muscle Innervation).
⒜이 근다발의 근섬유들에서는 몇 개의 신경근연접들이 보이고 있다. (LM× 253)
⒝신경근연접의 구조를 더욱 상세하게 보여주는 주사현미경 사진(SEM).
⒞주요 기능요소들과 신경근연접을 가로지르는 활동전위의 전달 단계들을 보여주는 도해 그림.
세사활주 이론은 수축시 근절에서 어떤 일이 일어나는지를 설명해준다. 이러한 관점을 갖고, 이제 수축과정을 좀더 상세히 살펴보기로 하자.
□ 골격근 활동의 제어(The Control of Skeletal Muscle Activity)
골격근섬유들은 오직 신경계의 통제하에서만 수축한다. 신경계와 골격근섬유들 사이의 전달은 신경근연접(neuromuscular junctions(NMJs)) 또는 근신경연접(myoneural junction)이라 하는 특화된 세포간 연결부에서 일어난다. 여러 가지 이 연접들이 그림 10-7a,b에 나타내져 있다.
신경근 연접(The Neuromuscular Junction)
각각의 골격근섬유들은 근섬유 길이의 중간에 있는 단 하나의 신경근연접에서 한 신경원(neuron)에 의해 통제된다. 그림 10-7c는 이 구조의 핵심적인 국면들을 요약하고 있다. 하나의 축색이 근다발막(perimysium) 안에서 분지하여 많은 가느다란 가지들을 형성한다. 이 각각의 가지들은 각기 부풀은 시냅스단추(synaptic knob)에서 끝을 맺는다. 시냅스단추의 세포질은 약자로 ACh라 하는 아세틸콜린(acetylcholine) 분자들로 채워진 소포들과 미토콘드리아를 수용하고 있다. 아세틸콜린은 신경전달물질(neurotransmitter)의 하나로, 신경원에서 유리되어 다른 세포들의 막의 성질을 변화시키는 화학물질이다. 시냅스단추에서의 아세틸콜린 유리는 골격근섬유의 수축을 격발하는 근초(sarcolemma)의 변화를 가져온다.
연접간극(synaptic cleft)이라 하는 좁은 공간이 시냅스단추 이와 마주보는 근초의 표면을 격리시킨다. 아세틸콜린이 부착하는 수용체를 내포하고 있는 이 표면을 운동종판(motor end plate)이라 한다. 운동종판에는 연접주름(junctional folds)이라 하는 깊은 주름들이 있는데, 이는 막의 표면적을 넓혀서 이용가능한 아세틸콜린 수용체의 수를 늘려준다. 아울러 연접간극과 근초는 아세틸콜린을 분해하는 효소인 아세틸콜린에스테라제(acetylcholinesterase (AChE) 또는 cholinesterase)를 수용하고 있다.
신경원이 근섬유를 자극하면 다음과 같은 일련의 단계로 수축과정이 일어난다.
1단계(Step 1): 활동전위의 도달(The Arrival of an Action Potential). ACh의 유리를 자극하는 것은 연접종말에 도달한 전기 임펄스, 즉 활동전위(action potential)이다. 활동전위는 축색을 따라 전파되는 막전위(transmembrane potential)의 급격한 변화이다.
2단계(Step 2): 아세틸콜린의 유리(The Release of ACh). 전기 임펄스가 연접종말에 도달하면, 막투과성이 변화되어 연접간극으로 ACh의 엑소시토시스(exocytosis)를 격발한다.
3단계(Step 3): 연접주름에서의 아세틴콜린 결합(ACh Binding at the Junctional Folds). ACh 분자들을 연접간극을 통해 확산되어 운동종판의 근초 표면의 ACh 수용체와 결합한다. ACh 결합이 일어나면, 근초의 나트륨이온 투과성이 바뀐다. 나트륨이온은 세포외액에서는 고농도로 수용되어 있지만, 세포내 농도는 매우 낮다. 막의 나트륨 투과성이 높아지면, 나트륨이온들 이 세포질내로 유입된다. 이 유입은 AChE가 수용체로부터 ACh를 제거할 때까지 계속된다.
4단계(Step 4): 근초의 활동전위 출현(Appearance of an Action Potential in the Sarcolemma). 나트륨이온들의 급격한 유입의 결과로 운동종판 주변 근초에 활동전위가 발생된다. 이 전기 임펄스는 막표면 전체로 퍼져나가 각각의 T세관(transverse tubule)들을 따라 전해진다. 따라서 신경종말에 도달한 활동전위가 근초의 활동전위 출현을 유도하게 된다.
흥분-수축 연결(Excitation-Contraction Coupling)
근초의 활동전위 발생과 근수축 시작 사이의 연결을 흥분-수축 연결(excitation- contraction coupling)이라 한다.
이 연결은 트라이어드(triads)에서 일어난다. 활동전위가 트라이어드에 도달하면, 이는 근형질세망(SR) 소조(cisternae)의 칼슘이온의 유리를 격발한다. 근형질세망의 칼슘이온 투과성 변화는 일시적으로, 단지 0.03초 정도만 지속된다. 그러나 근절(sarcomere) 내부와 주위의 칼슘이온 농도는 1/1000초 내에 휴지기 수준의 100배 정도에 이른다. 소조들은 두터운 세사와 가는 세사가 상호작용하는 곳인 겹친 구역(zones of overlap)에 위치하기 때문에, 칼슘 유리가 근절에 미치는 영향은 거의 즉각적이다. 세포질내 칼슘이온 농도 상승은 가는 세사를 따라 있는 활성부위(active site)를 노출시켜 수축을 시작시킨다. 이제 수축 주기(contraction cycle)가 시작된 것이다.
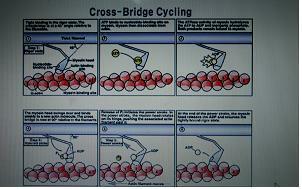
□ 수축 주기(The Contraction Cycle)
그림 10-8은 수축 과정중에 일어나는 분자수준의 사건들을 자세히 보여주고 있다. 과정의 최종 단계인 근절(sarcomere)내 각각의 연결다리(cross-bridge)에는 이미 활기찬 수축에 사용될 에너지가 충전되어 있다. 다리는 ATP를 분해할 수 있는 효소인 ATPase를 병합하고 있다. 수축 주기의 시작 시점에서, 각각의 연결다리들은 이미 ATP 한 분자를 분해해 수축과정에서 유리시킬 에너지를 저장하고 있다. 분해 산물인 ADP와 인산기(phosphate [PO43-])는 연결다리와 결합된 채로 남아있다.
수축 과정은 다음과 같이 연속된 다섯 단계가 관여한다.
1 단계(Step 1): 활성부위 노출(Active-site exposure). 근형질 안으로 들어온 칼슘이온들은 트로포닌(troponin)과 결합한다. 이 결합은 트로포닌 복합체와 액틴(actin)사이의 결합(bond)을 약화시킨다. 그렇게 되면 트로포닌 분자는 위치가 바뀌어 트로포미오신(tropomyosin) 분자를 활성부위에서 멀어지게 당긴다. (그림 10-8)
2 단계(Step 2): 연결다리 부착(Cross-bridge attachment). 활성부위가 노출되면, 미오신 연결다리가 활성부위와 결합한다.
3 단계(Step 3): 선회(Pivoting). 휴지 상태의 근절에서 각 연결다리들은 M선(M line)의 반대쪽을 향하고 있다. 이 위치에서 미오신 머리는 쥐덫의 스프링처럼 “장전되어(cocked)” 있다. 미오신 머리를 장전하는데 에너지가 소모되며, 이 에너지는 ATP를 ADP와 인산기로 분해하여 얻는다. 미오신 머리가 장전된 상태에서도 ADP와 인산기는 모두 여전히 미오신 머리에 결합되어 있다. 연결다리 부착이 일어난 뒤, 미오신 머리가 M선쪽으로 제껴지면서 저장되어 있던 에너지는 유리된다. 이것이 일어날 때 ADP와 인산기도 유리된다.
4 단계(Step 4): 연결다리 분리(Cross-bridge detachment). ATP 하나가 미오신 머리와 결합하게 되면, 액틴 분자의 활성부위와 미오신 머리 사이의 연결이 깨진다. 그렇게 되면 활성부위가 다시 노출되어 또 다른 연결다리와 상호작용할 수 있게 된다.
5 단계(Step 5): 미오신 활성화(Myosin activation). 결합에서 풀려난 미오신 머리가 ATP를 ADP와 인산기로 분해하면 미오신이 활성화된다. 이 과정에서 유리된 에너지는 미오신 머리를 재장전하는데 이용된다. 이제 전체 주기를 다시 반복할 수 있는 상태가 된 것이다. 만일 칼슘이온 농도가 여전히 상승되어 있고 ATP 비축량이 충분하다면, 각 미오신 머리는 이 주기를 초당 5회 정도 반복할 수 있다.
총괄적인 작용을 알아보기 위해, 여러분이 커다란 밧줄을 당기고 있다고 가정해 보자. 여러분은 미오신 머리이고, 밧줄은 가는 세사이다. 여러분이 앞으로 손을 뻗어 밧줄을 양손으로 움켜잡고 여러분 쪽으로 당긴다. 이 동작이 연결다리 부착과 선회에 해당한다. 여러분은 이제 밧줄을 놓고 다시 앞으로 손을 뻗어 밧줄을 움켜쥔다. 이 과정을 되풀이함에 따라 여러분은 밧줄을 차츰 당길 수 있게 된다.
이제는 여러 사람이 줄지어 서서 모두 같은 밧줄을 당기고 있는 상황을 생각해 보자. 각 사람마다 앞으로 손을 뻗어 밧줄을 움켜쥐고 당긴다. 다음에는 자기 앞에 있는 사람이 잡았다 놓은 곳을 손을 뻗어 움켜쥔다. 이같은 연속과정이 두터운 세사를 따라 있는 미오신 머리들이 함께 작용하여 한 가닥의 가는 세사를 근절 중앙쪽으로 당기는 방식과 일치한다.
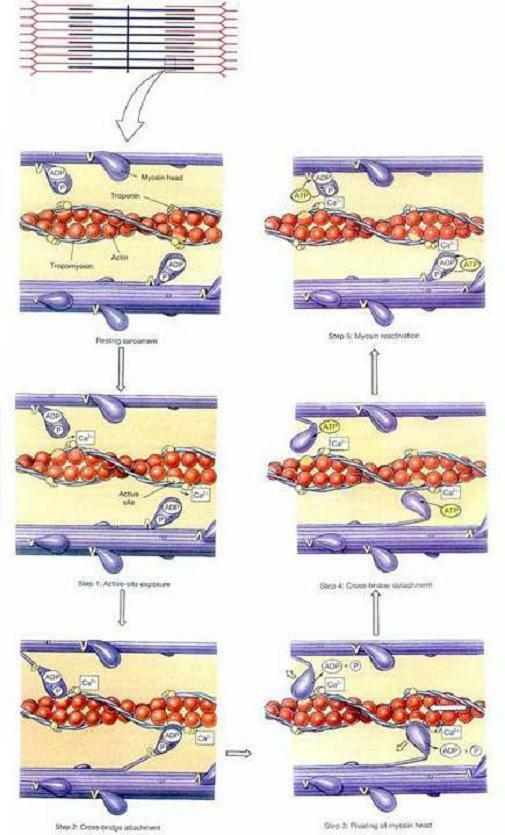
그림10-8. 수축과정의 분자수준의 사건들(Molecular Events of the Contraction Process)
3.움직임과 생각의 출현(이나스)
가. 운동성의 근원적 단계(myogenic stage)
“일반적인 뇌 구성 전개를 잘 예시하는 사례로 연골어류(상어)의 경우를 생각해보자. 상어의 배아는 산소가 통하는 알껍데기 속에서 발생한다. 발달 중인 배아의 조직에 산소가 골고루 분산되려면 알 안쪽에 있는 액체가 계속해서 움직여야 하므로, 배아는 사인곡선을 그리며 리듬있게 진동한다. 여기서 우리가 알게 되는 아주 중요한 점은 이 발달 단계에서는 운동이 신경계 활동에 의해 이루어지는 게 아니라는 사실이다.
근육조직으로 구성되고 리듬있는 운동을 하는 근육세포들은 아직 각각의 운동뉴런에 의해 자극될 필요가 없다! 그렇다면 근육이 어떻게 작동하는 것일까? 이 발달 단계에서 근육세포들은 모두 전기적으로 결합되어있다. ~이런 방식으로 결합되어 있으므로 한 근육세포의 수축을 일으키는 전기적 신호는 리듬 있는 진동이 완성된다. 이 운동은 순전히 근육세포 자체에서 생겨나므로, 이 사건을 운동성의 근원적 단계라고 한다.“
나. 신경적 운동성(neurogenic motricity)
“다음 발달 단계에서는 아주 중요한 기능적 변형이 일어난다. 척수가 목표 근육을 향해 여행하거나 이주하는 운동뉴런의 축색을 내보내기 시작하는 것이다. 이 시점에서 운동뉴런들도 전기긴장적으로 결합된다. 자라나는 축색이 목표 근육세포에 접촉하여 분포하기 시작하면 전기화학적 시냅스를 형성하고 동시에 근육세포는 전기적으로 결합된 상태를 마감한다. 이것이 근원적으로 유도된 운동성의 종말이다. 여기서 알 수 있는 건 진동운동을 만들어내는 능력이 근육세포에서 척수내부로 옮겨갔다는 사실이다. 근육덩어리의 운동성이 척수 뉴런 회로의 연결망과 본질적인 전기적 성질 안으로 유입되었다는 뜻이다. 이것이 신경적 운동성이라고 알려진 단계이다.”
다. 운동성의 내면화
“따라서 신경적 운동의 임피던스(impedance)는 근육의 성질과 밀접하게 대응된다. 결론은 동물의 외부 성질이 뇌 안에 내면화되기 시작했다는 것이다. 운동뉴런은 체계의 더 윗부분인 뇌간이 운동뉴런과 시냅스 연결을 시작할 때까지 전기긴장적 결합을 유지한다. 그때 운동뉴런의 전기긴장적 결합은 풀리지만 체계의 윗부분은 전기긴장적 결합을 유지한다. 이 발달 단계에서 운동뉴런은 전기긴장적 결합이 풀리게 되는 것 외에도 정해진 근육 집단의 활성화와 특정한 관계가 없는 신경계의 다른 부분들로부터 시냅스 입력을 받기 시작한다. 이렇게 추가된 입력은 동물의 전신 운동에 더 많이 관여하면서 전정계를 끌어들인다. 전정계한 운동성의 전신적 성질들에 관해 운동 신경망에게 정보를 주는 평형기관이다. 내가 똑바로 헤엄치고 있을까, 아니면 거꾸로 헤엄치고 있을까? 그것은 동물이 지구 중력이나 중력에 수직으로 운동하는 관성적 결과와 같이 자신의 몸보다 큰 좌표계를 고려하여 자신의 운동성을 조직하도록 돕는다. 그런 다음 대뇌화가 등장한다.”
a.단백질 사슬(액틴과 미오신)의 운동성
→단백질사슬(세포내 미세소관)들이 운동성을 극대화하기위해 선형으로 연결, 핵과 미토콘드리아는 찌그러든 형태로 존재, 소포체가 전체를 둘러싼다.
→다른 세포(전기적 pulse를 만들 수 있는 세포)의 막의 일부가 접촉, 전기적 충격을 연결시킨다.
→소포체(근형질세망)내의 Ca++이 단백질 사슬 사이로 방출, 세포내 액틴과 미오신의 활주가 가능해짐
b.세포의 미오닉 운동이 전기적 신경세포에 의해서 신경조절 형태로 운동이 조절
→ 척수전각에서의 하위운동신경원
“마음은 운동의 내면화이다” - 진화적으로 단세포의 미오닉한 운동성이 신경세포와 연접되면서 척수를 통해 중앙화, 내면화(척수를 통해 감각입력이 들어오고 운동계획, 운동출력이 나간다 - 감각운동이미지)
4.감각운동이미지 = 마음
"마음(mind) 혹은 마음상태(mindness state)란 자지 자각(self-awareness)을 포함해서 감각운동이미지(sensorymotor image)가 발생하는 전역적인(global) 뇌기능상태이다.
감각운동이미지(sensorymotor image)란 행동을 일으키는 하나의 구별되는 기능 상태를 만들어내는 데 관련되는 모든 감각 입력의 결합을 가리킨다.“
“예를 들어 등이 가렵다고 하자. 등은 볼 수 없는 곳에 있지만 가려움은 내부 ‘이미지’를 만들어서 복잡한 몸 안의 한 위치를 알려주고 취할 태도까지 알려준다. 긁어라! 그것이 감각운동 이미지이다. 감각운동 이미지의 발생은 단순한 입출력반응이나 반사가 아니다. 동물이 현재하고 있는 행위의 맥락안에서 일어나기 때문이다. 따라서 감각운동 이미지가 발생하고 운동 직전의 전운동(premotor)이 공식화될 때의 맥락은 내용만큼 중요하다.”
5.다양한 동물들의 운동 양태는 일률적이다. 
6.이 강의의 하나의 주제를 말한다면 세포(막)의 무한한 춤
- 세포의 다양한 무한 변형이 전자기적 현상을 드러낸 것이 우리의 의식
세포의 무한한 춤을 가능케 한 것
a.ATP
b.미토콘드리아와 엽록체
c.세포내 골격(액틴, 미오신)- 단세포의 운동이 가능해짐
-a,b,c.를 다 갖춘 것이 진핵세포
마음을 공부하려면 세포를 공부해야, 특히 연결을 위주로 하는 신경세포를 공부하자. 신경세포의 무수한 집합체가 곧 뇌이다.
7.운동과 의식의 유사성
근육운동은 필요에 의해서 (ex 컵을 잡는다) 개별 근육 대신 기능적 협동근육을 소환시켜 “가장 효과적으로 임무를 완수할 수 있는 협동 근육만을 고려함으로써 계산 부담을 더 낮춘다.” 운동은 줄곧 불연속으로, 박동으로 만들어지고 조절된다.
“유사한 방식으로 뇌는 실재 묘사기, 즉 의식 경험 생성기로 작용한다. 외부세계의 재구조물을 이음매 없이 흐르는 꿈과 같은 영화로 건네주기 위해서, 뇌는 영원히 앞을 내다보고 있어야 한다. 불연속적인 시점에 초점의 방향을 조정하면서, 불연속 시간 간격을 건너뛰는 와중에 그 모든 조각을 한데 모아야 한다.”
-의식도 monkey mind, 요동치는 토막 토막의 의식이 흐름을 이룬다. 반복되면서 무수하게 변형되는 소집되고 해산하는 방식이 유사하다.
“그런 의미에서 생각해 볼 때, 뇌는 어느 시점에서나 ‘오직 이 순간에 무엇을 아는 것이 중요한가(What-is-important-to-know-at-this-moment-only)'를 기초로 작용한다는 것이 분명해진다. 실제로 선택의 여지가 없다. 뇌는 다른 것을 할 시간이 없는 것이다!“
“그러므로 예측은 쉽게 해체되는 빠른 초점 재구성의 원동력임을 알 수 있다. 뇌가 무언가 알 혹은 사용할 필요 때문에 협동 근육을 소집하고 부린 다음 해산시키는 방식과, 의식 수준에서 초점이 다시 구성되는 방식은 동일하다. 운동과 인지를 위해 서로 다른 전략을 사용한다면, 그것은 이상한 뇌인 것이다.”
8.칼슘과 인의 위험한 정사
“세포 대 세포 의사소통에서의 위대한 발전은, 세포가 칼슘 이온 농도를 조절하기 위해 발달시킨 능력에서 나왔다. 칼슘은 주기율표에서 가장 반응성이 큰 원소들 중 하나로 극히 다루기 어려운 원소이다. ~칼슘은 돌을 만들지 않고서는 물에서 나트륨이나 칼륨처럼 높은 농도로 있을 수 없다. 돌이나 기타 결정은 생명과 정반대의 것이다. 그럼에도 불구하고 자연은 칼슘을 생명을 위한 필요조건으로 진화시켰고 그것을 매우 정확하게 조절하는 법까지 학습했다. 격렬하게 반응하는 이 원소가 어떻게 진핵 생명체의 구조 안으로 짜여 들어갔을까? 또 다른 원소인 인과 위험한 정사를 벌인 결과이다. 인은 진핵생명체에서 매우 중요하다. 근육 수축이나 신경세포 활동과 같이 에너지 소모가 많은 업무를 지원해야 하는 진핵 유기체에게는 연료 분자로부터 가장 질 좋은 가용에너지를 얻을 수단이 필요하다. 산화적 인산화 과정을 거치는 이 작업에는 산소가 필수적이다. 그러나 진핵 생명체가 산화적 인산화를 위해 인을 계속 싣고 다니려면 반응성이 높은 칼슘을 막아내는 법을 배워야 했다. 그렇지 않으면 칼슘은 세포에서 인을 훔쳐내어 결정화할 것이고 생명까지 훔쳐갈 것이다. ~인간이 불의 통제법을 배운 것처럼, 진핵생물은 칼슘의 통제법을 배웠다. 진화는 칼슘에 calmodulin이라는 단백질의 마구를 채워 극히 유용한 목적을 위해 이 원소의 반응성을 이용하기 시작했다. ~칼모듈린의 개발과 함께 칼슘/칼모듈린 착물은 아주 정교한 신호 체계로 중욯나 세포내 도구가 되었다. 보통 세포 내의 자유 칼슘 농도가 매우 낮은 덕분에 ‘2차 전령 역할’을 할 수 있었다. 이 역할은 정보 전달에 있어서 매우 중요하다. 효소 반응이 빠르고 국지적으로 유발되도록 조절하여 근육세포 수축, 축색연장, 시냅스 전달, 예정된 세포사망과 같은 많은 사건을 유도하기 때문이다. 진핵생물 진화에서 정점을 이루는 이 사건은 세포들이 상호 의사소통하는 공동체의 일부로 조직되기 위한 생물학적 필요조건들을 제공했다.”
세포내 단백질 + P = 단백질의 인산화(무생물과 같은 단백질이 인산화됨으로써 움직인다. 단백질의 에너지, 생화학 작용의 출발점)
칼슘의 농도는 세포내외의 차가 10만배 이상 차이가 난다. 세포내 농도가 낮다. 세포내 칼슘이온을 소포체 내에 가두어 두고 있으나 전기적 pulse를 받으면 Ca++가 세포내로 퍼지고 액틴과 미오신이 작동한다. 소포체 밖에 있어 PO4-와 만나면 세포가 죽는다. Ca++이 PO4-를 만나면 인산칼슘, 즉 뼈가 된다. 인산염과 칼슘을 분리시켜 놓는게 세포에 중요하다.
근형질 안으로 들어온 칼슘이온들은 트로포닌(troponin)과 결합한다. 이 결합은 트로포닌 복합체와 액틴(actin)사이의 결합(bond)을 약화시킨다. 그렇게 되면 트로포닌 분자는 위치가 바뀌어 트로포미오신(tropomyosin) 분자를 활성부위에서 멀어지게 당긴다. 활성부위가 노출되면, 미오신 연결다리가 활성부위와 결합한다. 휴지 상태의 근절에서 각 연결다리들은 M선(M line)의 반대쪽을 향하고 있다. 이 위치에서 미오신 머리는 쥐덫의 스프링처럼 “장전되어(cocked)” 있다. 미오신 머리를 장전하는데 에너지가 소모되며, 이 에너지는 ATP를 ADP와 인산기로 분해하여 얻는다. 미오신 머리가 장전된 상태에서도 ADP와 인산기는 모두 여전히 미오신 머리에 결합되어 있다. 연결다리 부착이 일어난 뒤, 미오신 머리가 M선쪽으로 제껴지면서 저장되어 있던 에너지는 유리된다. 이것이 일어날 때 ADP와 인산기도 유리된다. ATP 하나가 미오신 머리와 결합하게 되면, 액틴 분자의 활성부위와 미오신 머리 사이의 연결이 깨진다. 그렇게 되면 활성부위가 다시 노출되어 또 다른 연결다리와 상호작용할 수 있게 된다. 결합에서 풀려난 미오신 머리가 ATP를 ADP와 인산기로 분해하면 미오신이 활성화된다. 이 과정에서 유리된 에너지는 미오신 머리를 재장전하는데 이용된다. 이제 전체 주기를 다시 반복할 수 있는 상태가 된 것이다. 만일 칼슘이온 농도가 여전히 상승되어 있고 ATP 비축량이 충분하다면, 각 미오신 머리는 이 주기를 초당 5회 정도 반복할 수 있다.
9.ATP
생명현상에 있어 거의 초기단계에 만들어진 것이 ATP합성 메카니즘
ATP합성이야말로 의식의 출현에 가장 바탕이 되는 생명현상
ATP 나모머신이야말로 생명체의 가장 정교하고 먼저 생긴 머신
생명 현상의 가장 기본이 미토콘드리아고 미토콘드리아가 에너지 생성하는데 가장 기본이 양성자 pump이다. 생명 현상은 아직도 한번도 움직인 적이 없는 듯하다. 우리 몸을 이루는 가장 기본 메카니즘은 빅뱅이후 up quark 두 개와 down quark 하나가 양성자 하나를 만드는데 그것을 지금 그대로 사용하고 있다. 생명 현상에 있어 최소 5억년을 한 순간으로 봐야하고 좀 더 과장하면 생명현상은 35억년 동안 한 발자국도 변화하지 않았다. 생명체가 고등화하여 진화했다고 생각한다면 어처구니 없는 생각이다.
“우리 관점에서 생물들을 보면, 당신네 진핵생물들은 곧 그런 젠체하는 태도를 버릴 것이다. 당신네 두발 유인원들, 꼬리 잘린 나무땃쥐, 물기가 빠진 육기어류, 척추를 가진 벌레들, 혹스유전자를 늘린 해면동물들, 한꺼번에 무더기로 등장한 것들, 진핵생물들, 단조롭고 비좁은 영역에 옹기종기 모인 거의 구별되지 않은 존재들은 세균들의 표면에 있는 변덕스러운 거품에 다름 아니다. 당신들을 이루는 세포들 자체가 우리 세균들이 10억년 전에 발견한 낡은 기술들을 똑같이 재현하는 세균 군체들이기 때문이다. 우리는 당신들이 오기 전부터 여기에 있었고, 당신들이 떠난 뒤에도 여기에 남을 것이다.” - 조상 이야기
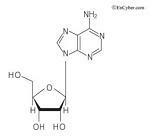
아데노신의 구조
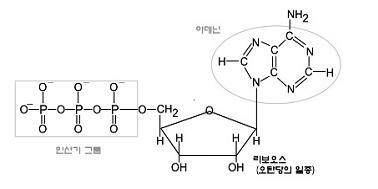
ATP의 구조