 제 16 강 감 정 의 뇌 2
제 16 강 감 정 의 뇌 2
1.brain은 거의 연합뉴런: 지연된 반응을 위해 연합뉴런이 많을 수밖에 없다.
감각뉴런 : 연합뉴런 : 운동뉴런 = 10 : 10만 : 1
(호모사피엔스의 운동은 신중하게 예측, 판단, 비교하여 계산된 운동)
시시덕이(떠벌이)는 재를 넘지만 샌님은 골로 빠진다.(속담)
-시시덕이는 복합계, 어디가도 communication 잘 하는 사람/ 샌님은 어려운 상황에 잘못된 판단을 많이 한다.
다마지오의 환자, 엘리옷이 안와전전두엽에 종양이 생긴 후 감정의 훼손으로 판단력이 저하되어 친구를 사기꾼을 사귀어서 파산하게 됨
→ 어떤 사람의 신뢰성 여부를 판단할 때 논리적으로 아는 게 아니라 적극적으로 느낌으로 안다.
비표준적인 다양한 입력에 대해 이성적으로 판단하는 게 아니라 즉각적으로 매순간 느낌을 통해 판단한다.
이성은 어쩌면 근거없는 토끼풀일 수 있지만 느낌이야말로 아주 고등한 명확한 근거가 있다.
전대상회, 복내측전전두엽 등 high level에서 정보를 수집하고 계산하지만 느낌의 표출은(감정의 신체적 표현-울고, 웃고 등등) 뇌간에서 기원한다.
감정이 세밀하고 느낌이 훌륭한 사람일수록 판단력이 뛰어나다..
2.하행운동신경
-진화순서
a.그물척수로(어류) :뇌간에 있다, 꼬리치는 것, 망상체(그물형성체)
b.전정척수로(양서류) :평형기능의 조절
육상동물로 진화 중력에 대해 균형을 잡는다.
균형은 시각, 평형감각, 체감각 중 두개 이상 동작해야 균형 잡힌다.
c.덮개척수로(시개척수로) :중뇌 상구, 동안근을 관장한다.
시각의 발달, 육지에서 목표를 향해가기 위해서
d.적핵(적색)척수로(중뇌 내부) : 사지 원위부의 운동조절 /
flexor굴근을 흥분시키고 extensor를 억제시키는 역할을 한다고 생각된다.
e.피질척수로(추체로, 피라미드로) : 영장류에 특히 인간이 발달.
직접 운동신경원을 통제, MI에서 내려오는 것, 사지말단의 정교한 운동(능숙한 운동)이 가능해짐.
외측섬유단(외측피질척수로) - MI 40%, 두정엽 30%, 전운동영역30%에서 하행하여 외측피질척수로를 따라 운동출력이 나오고 척수의 전각운동뉴런(하위운동신경원LMN)을 통해 사지로 출력
3.운동과 감각, 느낌
후섬유단에서 1차 체감각영역 : 섬세한 감각
↓ 느낌
피질척수로(추체로) : 능숙한 운동(손가락, 입술)
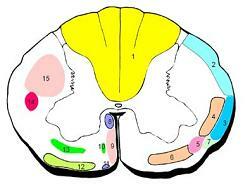
1.후섬유단(posterior funiculus, posterior white column)
2.후척수소뇌로(posterior spinocerebellar tract)
3.전척수소뇌로(anterior spinocerebellar tract)
.외측척수시상로(lateral spinothalamic tract)
5.척수덮개로(spinotectal tract)
6.전척수시상로(anterior spinothalamic tract) 7.척수올리브로(spino-olivary tract) 8.내측세로다발(medial longitudinal fasciculus) 9.전피질척수로(anterior corticospinal tract) 10.교뇌그물척수로(pontine reticulospinal tract) 11. 덮개척수로(tectospinal tract) 12. 전정척수로(vestibulospinal tract) 13. 수뇌그물척수로(medullary reticulospinal tract) 14. 적색척수로(rubrospinal tract) 15. 외측피질척수로(lateral corticospinal tract)
그림 5-10. 척수의 신경로(tracts of the spinal cord).
4.천재는 ‘머리’보다 ‘땀’이다.
천재 = 영감 1% + 땀 70% + 환경 29%(이인식의 ‘멋진 과학’)
양과 질에서 양이 먼저다. 양이 임계치를 넘으면 질로 바뀐다.
정보의 집적(신문이 유용)이 임계치를 넘어야 천재
“천재의 수수께끼에 도전한 인지과학자들은 천재나 범인, 모두 문제 해결방식이 동일한 과정을 밟는다는 사실을 밝혀냈다. 다시 말해 천재와 보통사람 사이의 지적 능력 차이는 질보다 양의 문제라는 것이다.” 이인식의 ‘멋진 과학’
예컨대 다산 정약용의 경우, 우리나라에서 가장 많은 저술을 남겼다. 그의 천재성은 다산 초당 11년, 강진 읍내 7년 유배생활동안 다져졌다. 다산초당 11년 동안 1000여권이상을 보았다. 엄청난 database에 접속해 있었다. 정보량과 정보를 연결시킬 수 있는 면밀한 관찰력, 고요한 내적 평정을 유배기간 얻어왔다.
5.후섬유단-내측섬유띠신경로
(Posterior White Column-Medial Lemniscal Pathway, Posteromedial System)
감각성분 : 분별촉각(discriminative touch)
진동감각(vibration and flutter sensation)
위치감각(position sensation)
수 용 기: 메르켈촉각소체(Merkel's disk)
마이스너촉각소체(Meissner's corpuscle)
크라우제종말망울(Krause's end bulb)
골지-마초니망울소체(Golgi-Mazzoni corpuscle)
파치니층판소체(Pacinian corpuscle)
모낭종말(hair follicle receptor)
루피니소체(Ruffini's corpuscle)
신경근방추(neuromuscular spindle)
골지힘줄기관(Golgi tendon organ)
제1차신경원: 척수신경절(spinal ganglion)
후근(posterior root)
후섬유단(posterior column, 후삭)
제2차신경원: 후섬유단핵(dorsal column nuclei, 후삭핵); 쐐기핵(nucleus cuneatus, 설상핵) 얇은핵(nucleus gracilis, 박속핵)
내궁상섬유(internal arcuate fiber)
내측섬유띠교차(lemniscal decussation, 내측모대교차)
내측섬유띠(medial lemniscus, 내측모대)
제3차신경원: 시상의 배쪽후외측핵 미부(VPLc of thalamus)
내섬유막(internal capsule, 내포)
방사관(corona radiata)
종 지: 중심후이랑(postcentral gyrus)의 일차감각영역(S I)
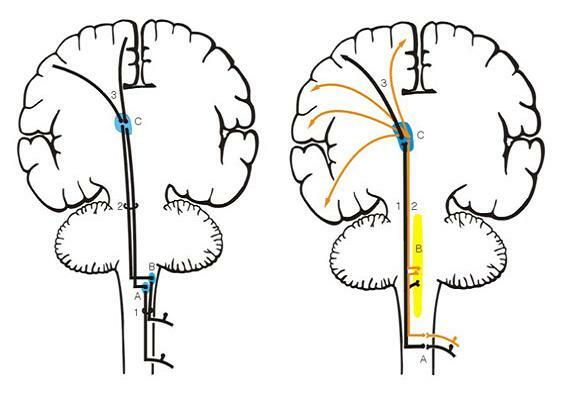
A. 얇은핵(nucleus gracilis, 박속핵) A. 척수 후각(posterior horn)
B. 쐐기핵(nucleus cuneatus, 설상속핵) B. 그물형성체(reticular formation)
C. 시상(thalamus, VPLc nucleus) C. 시상(thalamus, VPLc)
1. 후섬유단(posterior white column) 1. 신척수시상로(neospinothalamic tract)
2. 내측섬유띠(medial lemniscus) 2. 구척수시상로(paleospinothalamic tract)
3. 방사관(corona radiata) 3. 방사관(corona radiata)
그림 6-13. 척수에서 기원되는 상행피질계(ascending cortical system). 왼쪽은 후섬유단-내측섬유띠로(posterior white column-medial lemniscal pathway)이며 오른쪽은 척수시상로(spinothalamic tract)이다.
6.척수시상로 (Spinothalamic Tract, 전외측로 Anterolateral System)
척수시상로는 통증(pain)을 전달하기 때문에 임상적으로 매우 중요한 신경로이다. 온도감각(temperature)과 비분별성촉각(crude or light touch)도 전달한다고 알려져 있다. 척수의 전외측부에 있기 때문에 전외측로(anterolateral pathway or system)라고도 한다. 후섬유단-내측섬유띠신경로와 같이 시상을 거쳐 대뇌피질로 들어가 감각을 인지(지각 perception)하는 신경로이지만 후섬유단과는 달리 2차신경원이 척수 내에 있고, 척수에서 반대편으로 교차한 다음 상행한다. 대부분의 섬유가 시상으로만 가는 후섬유단-내측섬유띠신경로와는 달리 척수시상로는 뇌간의 그물형성체(reticular formation)로 많은 섬유를 보내는 것으로 알려져 있다. 과거에는 외측척수시상로(lateral spinothalamic tract)와 전척수시상로(anterior spinothalamic tract)를 구분하여, 외측척수시상로가 보다 외측에 위치하고 통각과 온도감각을 전달한다고 생각했고, 전척수시상로는 척수의 앞쪽에 위치하고 비분별성촉각을 전달한다고 생각했었으나, 현재에는 이러한 구분이 되는 뚜렷한 증거가 없다는 것이 밝혀졌기 때문에 이 두 신경로를 구분하지 않는 것이 일반적이다. 척수에서 이 두 신경로 사이에는 척수덮개로(spinotectal tract)가 위치해 있다
감각말단(감각수용기) - 무수축색이 피부 표면까지 나와있다.
미주신경, 부완핵PB과 고립핵NTS 쪽으로 가는 것은 insula를 통해 복내측전전두엽, 대상회 쪽으로 가면서 고차원적 느낌을 형성
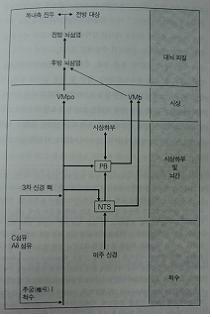
신체 내부 환경과 내부 장기의 신호를 뇌로 전달하는 데에서 필수적인 경로를 나타낸 것이다. 중요한 신호의 상당부분은 척수와 뇌간의 3차 신경핵을 통해서 전달된다. 말초신경섬유인 C형(전달 속도가 느린 가느다란 무수신경) 및 Aδ형 섬유가 전달해 준 정보는 척수의 모든 단계에서 추궁 I (척수의 회색질 뒤쪽의 뿔(horn) 부분과 3차 신경 핵의 꼬리 부분에 해당되는 영역)에서 중추 신경계로 보내진다. 이 정보들은 말 그대로 우리의 전신 구석구석에서 온 것으로 다양한 요소들과 관련되어 있다. 그 요소는 동맥의 평활근 수축 상태, 신체 국부의 혈액 흐름의 양, 국부의 체온, 각 조직에 중요한 손상을 일으킬 수 있는 화학물질의 존재, pH, 산소, 이산화탄소의 농도 등을 포함한다. 이 모든 정보들은 시상에 있는 전담 핵(VMpo)으로 전달되었다가 그 다음에는 후방 및 전방 뇌섬엽의 신경지도에 도달한다. 그러면 뇌섬엽은 복내측 전전두엽 피질이나 전방 대상 피질로 신호를 보낸다. 한편 정보는 시상으로 가는 길에 고립로 핵(nucleus tractus solitarius, NTS)에 들러 이곳에서도 이용 가능하게 된다. NTS는 미주신경(척수를 거쳐 전달되는, 대부분의 내부 장기들이 보내오는 신호들의 주요 경로)으로부터 신호를 받아서 완방핵(parabrachial nucleus, PB, 팔곁핵)과 시상하부로 보 대상 피질로 신호를 보낸다. 한편 정보는 시상으로 가는 길에 고립로 핵(nucleus tractus solitarius, NTS)에 들러 이곳에서도 이용 가능하게 된다. NTS는 미주신경(척수를 거쳐 전달되는, 대부분의 내부 장기들이 보내오는 신호들의 주요 경로)으로부터 신호를 받아서 완방핵(parabrachial nucleus, PB, 팔곁핵)과 시상하부로 보 대상 피질로 신호를 보낸다.
한편 PB와 NTS는 뇌섬엽으로도 신호를 보내는데, 이때는 다른 시상핵(VMb)을 통해서 전달한다. 흥미로운 것은 신체 운동이나 공간에서의 신체의 위치에 관련된 신호의 전달은 완전히 다른 경로를 통해 이루어진다는 것이다. 이러한 신호를 운반하는 말초신경섬유들(Aβ)은 두껍고 매우 빠른 속도로 신호를 전달한다는 점이다. 신체 운동에 관여하는 척수 및 3차 신경 핵, 그리고 시상 중계 핵과 최종적으로 신호가 도달하는 피질 영역(SI) 역시 차이를 보인다.
7.뇌신경중 미주신경은 내장에서 올라오고, 고립로핵은 미각과 관련
고립로핵 (nucleus solitarius)
연수의 상부를 지나는 수평단면에서 고립로핵은 내측전정핵의 앞쪽, 소세포그물핵의 뒤쪽, 삼차신경척수핵(spinal nucleus of trigeminal nerve)의 뒤쪽 내측에 위치해 있으며 제4뇌실에서 약간 떨어져 있다. 이 부분의 세포는 주위의 그물형성체의 세포와 모양이 비슷하기 때문에 경계가 불확실하며, 고립로(tractus solitarius)도 뚜렷하지 않다.
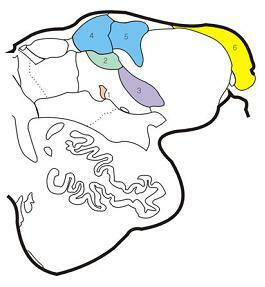
1.의문핵(ambiguus nucleus)
2.고립로핵(nucleus solitarius)
3.삼차신경척수핵 입쪽부분(pars oralis,
spinal tract nucleus of trigeminal nerve)
4.내측전정핵(medial vestibular nucleus)
5.하전정핵(inferior vestibular nucleus)
6.등쪽달팽이핵(dorsal cochlear nucleus)
그림 6-47.연수의 상부를 지나는 단면(upper level of medulla)의 뇌신경핵(cranial nerve nuclei).
고립로핵은 내장구심성핵(visceral afferernt nucleus)으로 구강, 비강, 인두, 식도, 위 등에서 들어오는 일반내장감각(general visceral afferent sensation)을 미주신경(X)의 하신경절(inferior ganglion)과 설인신경(IX)의 하신경절(inferior ganglion)에 있는 일차신경원을 통해 받아들이며, 혀와 후두덮개의 맛봉오리(taste bud)에서 감지하는 특수내장감각(special visceral afferent sensation)인 미각(taste)을 안면신경(VII)의 무릎신경절(geniculate ganglion)과 설인신경(IX)의 하신경절(inferior ganglion), 미주신경(X)의 하신경절에 있는 일차미각신경원을 통해 받아들인다. 고립로핵의 상부인 이 부분은 미각을 받아들이는 부분으로 미각핵(gustatory nucleus)이라고도 한다('섬유띠교차를 지나는 수평단면' 참조).
8.삼차신경 (trigeminal nerve, V)
삼차신경절이 얼굴전체에서 가장 중요하다.(턱, 혀 움직임과 얼굴에서의 체감각)
교뇌의 중간에서는 뇌간에서 나오는 뇌신경 중에서 가장 큰 신경인 삼차신경(trigeminal nerve, V)이 관찰된다. 체간의 촉각은 후섬유단을 통해, 안면의 촉각은 삼차신경
삼차신경은 얼굴의 대부분에서 일반감각을 받아들이는 감각근(sensory root)과 첫째아가미궁(first branchial arch)에서 기원되는 근육을 지배하는 운동근(motor root)으로 구성되어 있다.
성분(components): 삼차신경은 일반체구심성분(GSA)과 특수내장원심성분(SVE)의 두 가지 성분으로 구성된 혼합신경(mixed nerve)이다. 일반체구심성분(GSA)은 감각근을 통해 교뇌로 들어가며, 특수내장원심성분(SVE)은 운동근을 통해 근육을 지배한다.
일반체구심성분(GSA)의 세포체는 삼차신경절(trigeminal ganglion, 반월신경절 semilunar ganglion, 가서신경절 Gasserian ganglion)에 있으며 말초가지는 안신경(ophthalmic nerve), 상악신경(maxillary nerve), 하악신경(mandibular nerve)의 세 가지로 나누어져 얼굴, 이마, 비강 및 구강의 점막, 치아, 뇌경막 등에 분포한다.
삼차신경절신경원의 중심가지는 삼차신경의 감각근(sensory root)을 통해 교뇌로 들어가 삼차신경주감각핵(main sensory nucleus)과 삼차신경척수핵(spinal nucleus of trigeminal nerve)에 종지한다. 삼차신경척수핵으로 내려가는 중심가지섬유는 삼차신경척수로(spinal tract of trigeminal nerve)를 형성하며, 특히 통각과 온도감각을 전달하는 섬유는 삼차신경척수핵의 꼬리부분(pars oralis)에 종지한다.
일부 일반체구심섬유는 삼차신경절에 세포체가 있는 것이 아니라 교뇌 상부에서 중뇌의 상부까지 뻗어 있는 삼차신경중뇌핵(mesencephalic tract nucleus of trigeminal nerve)에 그 세포체가 있다. 삼차신경중뇌핵의 세포는 가단극신경원(pseudounipolar neuron)으로 중추신경계 내부에 있는 일차감각신경원이다. 이 핵 신경원의 말초돌기는 대부분 하악신경(mandibular division)을 통해 운동근의 축삭과 동반하여 저작근(masticatory muscle)의 신경근방추(neuromuscular spindle)와 치아, 치아주위막(periodontal ligament), 경구개(hard palate)에 분포하며, 중추돌기는 대부분 삼차신경운동핵(trigeminal motor nucleus)과 이어져 반사적으로 저작하는 힘을 조절하는 기전에 관여한다. 삼차신경중뇌핵의 일부 섬유는 소뇌와 이어져 있다.
삼차신경의 특수내장원심성분(SVE)은 삼차신경운동핵(trigeminal motor nucleus)에서 기원되며, 운동근(motor root)을 형성하여 첫째아가미궁(first branchial arch)에서 기원된 저작근(muscles of expression)-교근(masseter muscle), 내측날개근(medial pterygoid muscle, 내측익돌근), 외측날개근(lateral pterygoid muscle, 외측익돌근), 측두근(temporalis muscle)-과 하악설골근(mylohyoid muscle), 이복근(digastricus muscle)의 전복(anterior belly), 구개긴장근(tensor veli palatini muscle), 고막긴장근(tensor tympani muscle)을 지배한다.
분포: 삼차신경은 교뇌의 중간부분에서 외측으로 나오며 큰 감각근(sensory root)과 작은 운동근(motor root)으로 구성되어 있다. 운동근은 감각근의 내측 앞쪽에 있다. 이 신경의 감각신경절인 삼차신경절(trigeminal ganglion)은 반달모양이며 크기는 1 cm x 2 cm 정도이다. 신경절의 오목한 쪽으로는 중추돌기가 나와 감각근을 형성하며, 볼록한 쪽으로는 말초돌기가 세 개의 신경섬유다발을 형성하여 바깥쪽으로 나온다
 1.안신경(opthalmic nerve, V1)
1.안신경(opthalmic nerve, V1)
2.상악신경(maxillary nerve, V2)
3.하악신경(mandibular nerve, V3)
4.삼차신경절(trigeminal ganglion)
5.전두신경(frontal nerve)
6.비섬모체신경(nasociliary nerve)
7.누선신경(lacrimal nerve),
8.관골신경(zygomatic nerve)
9.날개구개신경절(pterygopalatine ganglion)
10.후상치조가지(superior posterior
alveolar branches)
11.구개신경(palatine nerve)
12.볼신경(buccal nerve, 협신경)
13.혀신경(lingual nerve, 설신경) 14.하치조신경(inferior alveolar nerve)
15.귓바퀴측두신경(auriculotemporal nerve 이개측두신경)
16. 하악신경절(submandibular ganglion)
그림 6-72.삼차신경(trigeminal nerve)의 가지(branches)와 삼차신경이 분포하는 얼굴의 영역.
이 중에서 안신경(ophthalmic nerve)은 상안와틈새(superior orbital fissure)를 통해 두개강의 바깥쪽으로 나오며, 이마(forehead), 위쪽 눈꺼풀(upper eyelid) 및 코의 피부(skin)와 안구(eyeball), 결막(conjunctiva), 누선(lacrimal gland) 및 비강(nasal cavity)과 부비동(paranasal sinus)의 점막(mucous membrane)에 분포한다.
상악신경(maxillary nerve)은 안신경과 하악신경 사이에서 나오며 원형공(foramen rotundum)을 통해 두개강 바깥으로 나온다. 얼굴의 중간부분, 아래 눈꺼풀, 코의 외측, 윗입술(upper lip)의 피부와 연구개(soft palate), 경구개(hard palate), 편도(tonsil), 위쪽 잇몸(upper gum)과 치아(upper teeth) 및 상악동(maxillary sinus)의 점막에 분포한다.
하악신경(mandibular nerve)은 삼차신경의 세 신경다발 중 가장 크며 운동근의 특수내장원심(SVA)성분도 포함되어 있는 혼합신경(mixed nerve)이다. 타원공(foramen ovale)을 통해 두개강 바깥으로 나온다. 감각섬유는 얼굴의 아래쪽, 뺨, 귀의 앞쪽 및 그 위쪽의 피부와 뺨(cheek), 혀, 구강바닥(floor of mouth)의 점막, 아래쪽 치아와 잇몸에 분포한다. 운동섬유는 저작근(muscles of expression)과 하악설골근(mylohyoid muscle), 이복근(digastricus muscle)의 전복(anterior belly), 구개긴장근(tensor veli palatini muscle), 고막긴장근(tensor tympani muscle)을 지배한다.
이차연결: 삼차신경핵감각핵은 시상과 소뇌로 원심섬유를 보내며, 여러 뇌신경핵과 반사궁으로 연결되어 있다.
삼차신경감각핵에서 나오는 이차삼차섬유(secondary trigeminal fiber)는 각 신경원이 위치한 부분에서 반대쪽으로 교차하여 상행하는 신경로인 배쪽삼차시상로(ventral trigeminothalamic tract)와 동측으로 상행하는 등쪽삼차시상로(dorsal trigeminothalamic tract)를 형성하며, 시상의 배쪽후내측핵(ventral posteromedial nucleus, VPM)에 종지한다.
삼차신경척수핵의 중간부분(pars interpolaris)과 입쪽부분(pars oralis)에서 나오는 이차섬유는 삼차소뇌섬유(trigeminocerebellar fiber)를 형성하여 소뇌로 투사되며, 삼차신경중뇌핵의 중추돌기 역시 소뇌로 투사된다고 알려져 있다.
일부 삼차신경감각핵의 섬유는 뇌간의 운동신경핵과 연결되어 있어 여러 가지 반사작용에 관여한다. 삼차신경중뇌핵의 중추돌기는 삼차신경운동핵과 연결되어 반사적으로 씹을 때의 힘을 조절해주는 역할을 하며, 단시냅스성반사인 하악반사(jaw jerk, 교근반사 masseter reflex)의 경로도 된다.
일부 섬유는 안면신경핵과 연결되어 각막반사(corneal reflex)를 일으킨다. 각막을 자극했을 때 양쪽 눈을 감게 되는 이 반사는 안신경-삼차신경절-삼차신경감각근을 거쳐 들어온 섬유가 삼차신경감각핵으로 들어가고, 감각핵의 이차감각섬유는 양쪽 안면신경핵과 연결되어 있기 때문에 일어난다. 한쪽의 안신경이 손상되면 이 반사가 일어나지 않으나, 반사궁의 원심부분은 안면신경(VII)이므로 정상이기 때문에 반대쪽의 각막을 자극하면 양쪽 눈을 정상적으로 감는다.
재채기반사(sneezing reflex)와 구토반사(vomiting reflex)는 이차삼차섬유가 의문핵(nucleus ambiguus)과 주위의 그물형성체로 연결되어 일어난다. 침분비반사(salivary reflex)는 이차삼차섬유가 하타액핵(inferior salivatory nucleus)으로 연결되어 이루어진다.
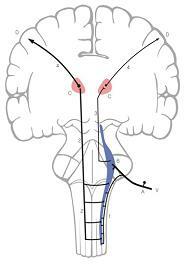
A.삼차신경절(trigeminal ganglion)
B.삼차신경감각핵(trigeminal sensory nucleus)
C.시상(ventral posteromedial nucleus, VPM)
D.대뇌피질(cerebral cortex)
1.삼차신경척수로(spinal tract of trigeminal nerve)
2.배쪽삼차시상로(ventral trigeminothalamic tract)
3.등쪽삼차시상로(dorsal trigeminothalamic tract)
4. 방사관(corona radiata)
V. 삼차신경(trigeminal neve)
그림 6-14. 삼차시상로(trigeminothalamic tract)의 경로. 삼차시상로는 얼굴의 일반감각(general sensation)을 전달하는 상행피질계(ascending cortical system)의 전도로이다.
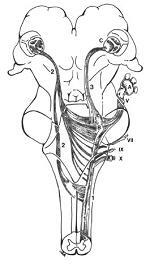
A.삼차신경절(trigeminal or semilunar ganglion)
B.삼차신경감각핵(trigeminal sensory nucleus)
C.시상(thalamus)
1.삼차신경척수로(spinal tract of trigeminal nerve)
2.배쪽삼차시상로(ventral trigeminothalamic tract)
3.등쪽삼차시상로(dorsal trigeminothalamic tract)
V.삼차신경(trigeminal neve)
그림 6-15. 입체적으로 나타낸 뇌간의 삼차시상로(trigeminothalamic tract).
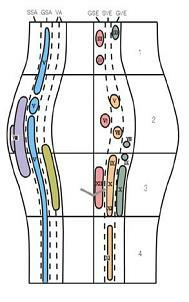
1. 중뇌(midbrain)
2. 교뇌(pons, 교, 뇌교)
3. 연수 상부-개방연수(open medulla)
4. 연수 하부-폐쇄연수(closed medulla)
GSE: 일반체원심성분 / SVE: 특수내장원심성분
GVE: 일반내장원심성분 / VA: 내장구심성분(GVA와 SVA)
GSA: 일반체구심성분 / SSA: 특수체구심성분
그림 6-4. 뇌간에서 뇌신경(cranial nerve)의 성분(components)과 위치. 신경의 성분에 따라 세로방향으로 긴 세포기둥을 이루고 있다. 중앙부에서 왼쪽은 감각핵(sensory niuclei)을 표시하였고, 오른쪽은 운동핵(motor nuclei)을 표시하였다.

1. 에딩거-베스트팔핵(Edinger-Westphal nucleus)
2. 동안신경핵(oculomotor nucleus)
3. 도르래신경핵(trochlear nucleus, 활차신경핵)
4. 삼차신경운동핵(trigeminal motor nucleus)
5. 외전신경핵(abducens nucleus)
6. 안면신경핵(facial motor nucleus)
7. 상타액핵(superior salivatory nucleus, 위침분비핵)
8. 하타액핵(inferior salivatory nucleus, 아래침분비핵) 9. 설하신경핵(hypoglossal nucleus)
10. 미주신경등쪽핵(dorsal motor nucleus of vagus nerve) 11. 의문핵(ambiguus nucleus, 의핵)
12. 척수부신경핵(spinal nucleus of accessory nerve)
그림 6-5. 뇌간의 원심성핵(efferent nuclei)과 뇌신경(cranial nerve)의 위치.
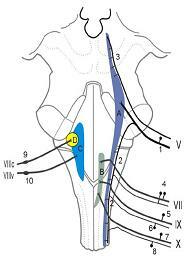
1. 삼차신경감각핵(trigeminal sensory nucleus)
2. 고립로핵(solitary tract nucleus)
3. 달팽이핵(cochlear nucleus)
4. 전정신경핵복합체(vestibular nuclear complex)
5. 삼차신경척수로(spinal tract of trigeminal nerve)
6. 삼차신경중뇌로(mesencephalic tract of trigeminal
nerve) 7. 삼차신경절(trigeminal ganglion)
8. 무릎신경절(geniculate ganglion)
9. 설인신경 하신경절(inferior ganglion of
glossopharyngeal nerve)
10. 설인신경 상신경절(superior ganglion of
glossopharyngeal nerve)
11. 미주신경 하신경절(inferior ganglion of vagus nerve)
12. 미주신경 상신경절(superior ganglion of vagus nerve)
그림 6-6. 뇌간의 구심성핵(afferent nuclei)과 뇌신경(cranial nerve)의 위치.

1. 에딩거-베스트팔핵(Edinger-Westphal nucleus)
2. 동안신경핵(oculomotor nucleus)
3. 도르래신경핵(trochlear nucleus, 활차신경핵)
4. 삼차신경운동핵(trigeminal motor nucleus)
5. 외전신경핵(abducens nucleus)
6. 안면신경핵(facial motor nucleus)
7. 상타액핵(superior salivatory nucleus)
8. 하타액핵(inferior salivatory nucleus)
9. 설하신경핵(hypoglossal nucleus)
10. 미주신경등쪽핵(dorsal motor nucleus of vagus nerve) 11. 의문핵(ambiguus nucleus, 의핵)
12. 척수부신경핵(spinal nucleus of accessory nerve)
13. 삼차신경중뇌핵(mesencephalic nucleus of trigeminal nerve) 14. 삼차신경주감각핵(main sensory nucleus of trigeminal nucleus) / 15. 삼차신경척수핵(spinal nucleus of trigeminal nucleus) /16. 달팽이핵(cochlear nucleus, 와우핵) / 17. 전정신경핵(vestibular nucleus) / 18. 고립로핵(solitary tract nucleus)
그림 6-7. 입체적으로 나타낸 뇌간의 뇌신경핵(cranial nerve nuclei)과 뇌신경(cranial nerve)의 위치
1. 적색핵(red nucleus)
2. 설하신경주위핵(peri-hypoglossal nucleus)
3. 전정신경핵(vestibular nucleus)
4. 하올리브핵복합체(inferior olivary nuclear complex)
5. 부쐐기핵(accessory cuneate nucleus 부설상속핵)
그림 6-8. 소뇌와 연결된 뇌간의 핵
(전소뇌핵 precerebellar nuclei).
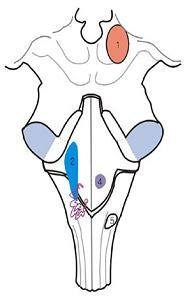
9.안면신경
A.안면신경마비(facial palsy)
B.A+타액분비(salivation) 감소
C.A+B+청각과민(hyperacusis)
D.A+B+C+눈물분비
(lacrimal secretion)감소
E.중추형 안면신경마비
(central type facial palsy)
1.안면신경핵(facial motor nucleus, SVE)
2.상타액핵(superior salivatory nucleus, GVE)
3.고립핵(solitary nucleus, VA) 4.삼차신경감각핵 (trigeminal sensory nucleus, GSA)
그림 6-61.안면신경(facial nerve)의 경로와 손상시 나타나는 증상.
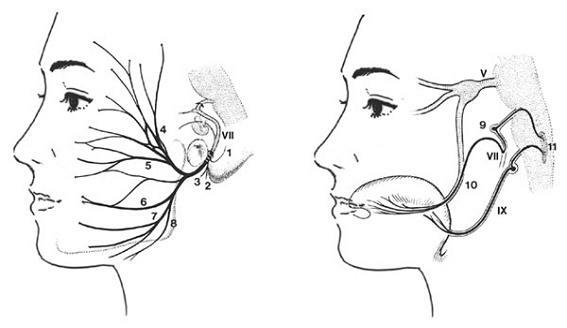
1.후귓바퀴신경(posterior auricular nerve, 후이개신경) 2.이복근가지(digastric branch, 이복지) 3.경상설골근가지(stylohyoid branch) 4.측두가지(temporal branch, 측두지)
5.관골가지(zygomatic branch, 관골지) 6.볼가지(buccal branch, 협지)
7.하악변연가지(marginal mandibular branch) 8.경부가지(cervical branch, 경지)
9.무릎신경절(geniculate ganglion) 10.혀신경(lingual nerve)
11.고립로핵(nucleus tractus solitarius) 12.설인신경 하신경절(inferior ganglion)
그림 6-60.안면신경 특수내장원심성분(SVE)의 종말가지인 말초운동가지(peripheral motor fiber)의 분포(왼쪽) 및 혀에 분포하는 특수내장구심성분(SVE, 미각 taste sensation) 신경의 분포경로(오른쪽).
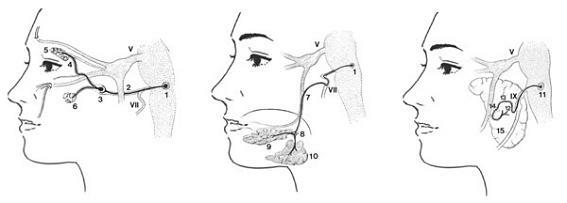
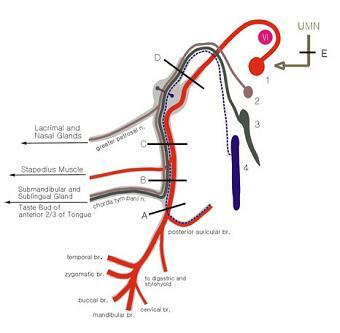
1.상타액핵(superior salivatory nucleus) 2.큰암석신경(greater petrosal nerve) 3.날개구개신경절(pterygopalatine ganglion) 4.누선신경(lacrimal nerve) 5.누선(lacrimal gland, 눈물샘) 6.비선(nasal glands)
7.고실끈신경(chorda tympani nerve) 8.악하신경절(submandibular ganglion) 9.설하선(sublingual gland, 혀밑샘) 10.악하선(submandibular gland, 턱밑샘) 11.하타액핵(inferior salivatory nucleus) 12.고실신경(tympanic nerve) 13.작은암석신경(lesser petrosal nerve) 14.이신경절(otic ganglion, 귀신경절) 15.이하선(submandibular gland, 귀밑샘)
그림 6-59.얼굴의 분비선에 분포하는 부교감신경섬유(parasympathetic fiber). 안면신경(facial nerve, VII)과 설인신경(glossopharyngeal nerve, IX) 일반내장원심성분(GVE)의 분포.
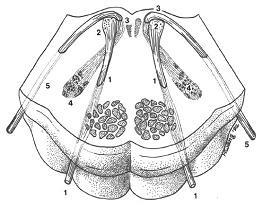
1.외전신경(abducens nerve)
2.외전신경핵(abducens nucleus)
3.안면신경무릎(genu of facial nerve, 안면신경슬)
4.안면신경핵(facial motor nucleus)
5.안면신경(facial nerve)
그림 6-58.안면신경(facial nerve, VII)과 외전신경(abducens nerve, VI)의 관계를 입체적으로 나타낸 그림. 안면신경은 안면신경핵(facial motor nucleus)을 나와 등쪽 내측으로 진행하여 외전신경핵(abducens nerve)의 등쪽을 돌아(안면신경무릎 genu of facial nerve) 외측으로 뇌간을 나오고, 외전신경은 외전신경핵에서 곧바로 내측으로 뇌간을 나온다. 능형오목(rhomboid fossa)의 안면신경구(facial colliculus)는 안면신경무릎의 바로 바깥쪽에 있다. 따라서 안면신경구의 심부에는 안면신경핵이 아니라 외전신경핵이 위치하게 된다.
10.유체이탈('out of body' experiences)
유체이탈은 누구나 가능한 뇌경험
체감각은 환각을 쉽게 일으킨다.(시각, 청각의 환각은 병적이지만 체감각의 환각은 보편적)
시각, 청각은 잘 간섭 받기 힘들다.
느낌은 다른 뇌영역에 의해 쉽게 간섭받는다. 공감, 감정이입이 가능해짐.
연합체감각영역에 기억을 불러일으켜 감각과 matching을 시키는데 연합체감각영역에 문제가 있으면 인식불능증이 생긴다.
insula와 연합체감각영역에서 감정과 느낌을 만든다.
체감각의 환상은 본질적이다. 타인의 마음, 타인의 느낌을 읽을 수 있다.
다마지오 실험 예 - 인물 사진의 표정을 보고 그 사진 속 인물의 느낌을 읽어내는 실험, 두 종류의 환자층에서 느낌을 읽어내는 데 실패, 하나는 2차시각영역(시각정보를 모아 전체 그림을 만든다)의 손상이 있는 환자 또 하나는 insula 손상
1차시각에서는 바깥세상을 등형으로 있는 그대로 mapping, 2차시각영역 이상에서는 왜곡
시각은 구성적이다. 시각은 움직임에 대해 민감, 상구는 움직이는 물체에 detect
brain은 구성적이다. 의식 level에 올라오는 것은 계산결과만이 올라온다.
뇌활동 중 95%는 의식되지 않는다 :
의식화 되지 않는 부분 중 상당수가 소뇌에서 우리 몸 각 근육에서 오는 정보를 종합하고 균형을 잡는 것 → 거의 의식되지 않는다.
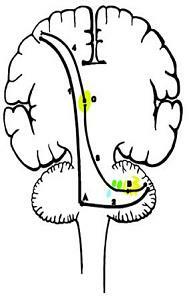
A. 교뇌핵(pontine nuclei)
B. 치아핵(dentate nucleus)
C. 시상(thalamus, VPLo, VLc)
D. 대뇌피질(cerebral cortex)
1. 피질교뇌섬유(corticopontine fiber)
2. 교뇌소뇌로(pontocerebellar fiber)
3. 치아시상섬유(dentatothalamic tract)
4. 시상피질섬유(thalamocortical fiber)
그림 7-17. 신소뇌(neocerebellum, 교뇌소뇌 pontocerebellum)의 연결.
대뇌피질이 운동에 필요한 정보를 소뇌피질에서 얻기 위한 폐loop
대뇌피질→뇌교핵→소뇌피질→소뇌심부핵→시상→대뇌피질SI
척추를 통해 올라오는 고유감각의 결과를 대뇌피질에서 교뇌를 통해 소뇌에 요구