
제14강 시각과 뇌
1.눈의 구조
눈의 벽은 셋으로 구별되는 층, 즉 세 가지 막-바깥쪽의 섬유막(fibrous tunic), 중간의 혈관막(vascular tunic), 그리고 안쪽의 신경막(neural tunic)을 수용하고 있다. 안구는 속이 비어있으며 안은 두 개의 강(cavity)으로 나뉠 수 있다. 큰 후강(posterior cavity)은 초자방(vitreous chamber)이라고도 부르는데, 교질성의 초자체(vitreous body)를 수용하고 있기 때문이다. 작은 전강(anterior cavity)은 다시 두 개의 방(chamber)-전안방과 후안방-으로 세분된다. 눈의 형태의 고정은 초자체에 의해 또 전강을 채우고 있는 투명한 방수(aqueous humor)에 의해 일부 이루어진다.
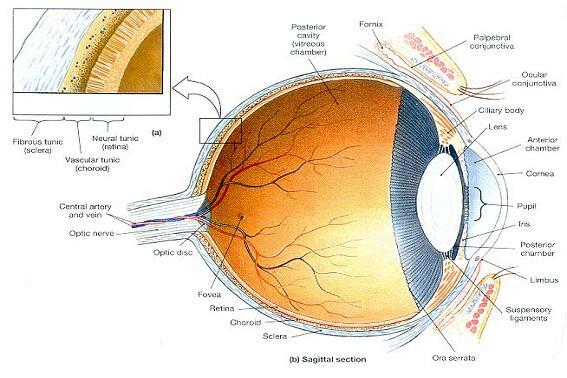
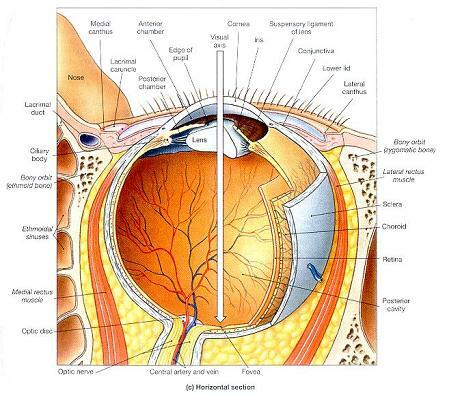
그림 17-8. 눈의 단면 해부학 (Sectional Anatomy of the eye).
⒜ 눈의 세 층, 즉 세 막.
⒝ 왼쪽 눈의 시상단면에서 보이는 주요한 해부학적 지표와 모습들.
⒞ 왼쪽 눈의 횡단면에서 보이는 지표와 모습들.
섬유막(The Fibrous Tunic)
눈의 가장 바깥층인 섬유막은 공막(sclera)과 각막(cornea)으로 구성된다. 섬유막은 ⑴역학적 지지와 어느 정도의 물리적 보호를 제공하고, ⑵외재안근의 부착부위로 제공되며, ⑶초점을 맞추는 과정을 돕는 구조물들을 수용하고 있다.
안구 표면의 대부분은 공막(sclera)에 의해 덮여져 있다. 공막, 즉 “눈의 흰자위(white of the eye)”는 교원섬유와 탄성섬유를 모두 수용하고 있는 치밀결합조직으로 구성되어 있다. 이 층은 시신경의 출구 부근인 눈의 뒤쪽이 가장 두텁고, 전면이 가장 얇다. 여섯 개의 외래안근(extrinsic eye muscles)이 공막에 종지하여, 자신들의 교원섬유와 이 바깥막의 교원섬유를 섞고 있다.
공막의 표면은 공막을 관통해 내부 구조물에 도달하는 작은 혈관과 신경들을 수용하고 있다. 안결막 밑에 있는 작은 혈관들로 된 혈관망은 보통 공막에 뚜렷한 색깔이 나타날 정도로는 혈액을 공급하지 않으나, 자세히 관찰하면 혈관들이 하얀 교원섬유 바탕에 있는 붉은 선들로서 보인다.
투명한 각막은 구조상 공막에 연속되어 있으며, 각막윤부(limbus)가 이 둘 사이의 경계이다. 각막상피 밑으로, 각막은 여러 층의 교원섬유를 수용하고 있는 치밀한 간질로 주로 구성되어 있다. 이 섬유들은 빛의 통과를 방해하지 않도록 일련의 층으로 편성되어 있다. 각막에는 혈관이 없고, 천층의 상피세포들은 반드시 노출면(free surface) 위로 흐르는 눈물로부터 산소와 양분을 얻어야만 한다.
각막에는 많은 수의 자유신경종말이 있으며, 각막은 눈에서 가장 민감한 부분이다. 광수용기를 포함해 눈의 나머지 부분은 완전히 정상일지라도, 각막 손상 하나만으로 실명을 일으킬 수도 있다. 불행하게도, 각막은 매우 제한된 복구 능력을 갖고 있다. 따라서 각막이 손상되었다면 심각한 시력 상실이 생기지 않게 반드시 즉시 치료를 받아야 한다. 각막의 상흔이 생긴 뒤(after corneal scarring) 시력을 복구하려면, 대부분 각막이식술(corneal transplant)을 통한 각막 교체가 필요하다. 각막교체는 아마도 가장 흔한 유형의 이식수술일 것이다. 이 이식은 연관이 없는 사람들 사이에서도 시술이 가능한데, 이형 조직(foreign tissues)을 공격하는 백혈구들을 이 부위로 운반해줄 혈관이 없기 때문이다. 각막이식편(corneal grafts)은 질병이나 사고로 사망한 공여자의 눈에서 얻을 수 있는데, 가장 좋은 결과를 위해서는 반드시 공여자가 사망한 지 5시간 이내에 조직을 떼어내야만 한다.
혈관막(The Vascular Tunic; 포도막(Uvea))
혈관막(vascular tunic), 즉 포도막(uvea)은 수많은 혈관과 림프관 및 내재안근들을 수용하고 있다. 이 층은 ⑴ 눈의 조직으로 공급되는 혈관과 림프관의 통로를 제공하고, ⑵ 눈으로 들어가는 빛의 양을 조절하며, ⑶ 안구 안을 순환하는 방수(aqueous humor)를 분비하고 흡수하며, ⑷ 초점을 맞추는 과정의 핵심 부분인 수정체의 형태를 조절하는 기능을 한다. 혈관막은 홍채(iris), 모양체(ciliary body), 그리고 맥락막(choroid)을 포함하고 있다.
홍채(The Iris)
투명한 각막면을 통해 볼 수 있는 홍채는 혈관, 색소세포 및 두 층의 평활근섬유층을 수용하고 있다. 이 근육들이 수축하면 홍채 중심 구멍, 즉 동공(pupil)의 직경이 변하게 된다. 한 무리의 평활근섬유들은 동공을 둘러싸는 일련의 동심원을 형성한다. 이 동공축소근(pupillary constrictor muscles)이 수축하면, 동공의 직경이 감소한다. 다른 무리의 평활근은 동공의 가장자리로부터 방사상으로 뻗어나가고 있다. 이 동공확대근(pupillary dilator muscles)의 수축은 동공을 확대시킨다. 두 무리의 근육 모두가 자율신경계에 의해 제어된다. 예를 들어, 부교감 활성화는 밝은 빛에 반응하여 동공 축소를 일으키고, 교감 활성화는 어둠침침한 빛에 반응하여 동공의 확대를 일으킨다(동공반사(pupillary reflex)).
홍채의 몸체(body)는 결합조직으로 구성되어 있으며, 그 후면은 색소세포를 내포하는 상피에 의해 덮여 있다. 색소세포는 홍채의 결합조직에도 존재할 수 있고 전면을 덮고 있는 상피내에 존재하기도 한다. 눈 색(eye color)은 색소세포의 밀도와 분포에 의해 결정된다. 홍채 결합조직에 색소세포가 없다면, 빛이 홍채를 통과해 홍채의 후면을 덮고 있는 색소상피(pigmented epithelium)의 내면에서 반사된다. 그러면 눈이 파랗게 보인다. 회색이나 갈색 및 검은 색의 눈을 가진 사람들은 홍체의 몸체와 표면에 훨씬 많은 수의 색소세포를 가지고 있다. 백변증인 사람(human albino)의 눈은 매우 옅은 회색 또는 청회색을 띤다.
모양체(The Ciliary body)
홍채의 바깥둘레는 모양체의 앞부분에 부착되어 있다. 모양체(ciliary body)는 각막과 공막 사이의 연접에서 시작된다. 모양체는 뒤로 시각수용기를 내포하고 있는 신경망막(neural retina)의 톱니같은 전연인 거상연(ora serrata)까지 뻗어있다. 모양체의 몸체는 눈의 안쪽으로 돌출되어 있는 가락지 모양의 근육인 모양체근(ciliary muscle)으로 구성되어 있다. 상피는 모양체 돌기(cilliary process)라 하는 수많은 주름들로 마무리된다. 이 돌기들에는 수정체의 현수인대(suspensory ligaments)가 부착한다. 이 결합조직섬유들이 수정체를 중심은 동공에 있으면서 홍체의 뒤에서 유지되게 한다. 그 결과, 동공을 통해 광수용기로 향하는 모든 빛은 수정체를 통과하게 된다.
맥락막(The Choroid)
맥락막(choroid)은 거상연 뒤에서 섬유막과 신경막을 분리시켜준다.(그림 17-8) 이것은 공막에 의해 덮여 있고 망막의 가장 바깥층에 부착되어 있다. 맥락막은 산소와 양분을 망막으로 운반하는 대량의 모세혈관망을 수용하고 있다. 아울러, 맥락막은 흩어져 있는 멜라닌세포들을 수용하고 있는데, 이들은 특히 공막 부근에 많이 있다.
2.신경막(The Neural Tunic)
①시신경층(원추세포 500만개, 간상세포 1억개) / Horizontal cell / ②Bipolar cell / Amacrine cell / ③신경절세포 - 신경절세포가 모여 다발을 이루어 나가는 곳이 맹점optic disc → ④LGN(lateral geniculate nucleus, 크릭이 만년에 20년간 뇌연구를 했는데 연구소 첫 부임 때 한 것이 외측슬상체에 색을 표했다- 시각시스템을 통해 의식구명하려고 함) 시각시스템의 중계기지 → VI(⑤단순세포-선, 점 / ⑥복합세포-선의 이동 / ⑦초복합세포-모서리, 가진 부위의 움직임) → V7(후두 피질의 60%가 시각을 process)
그림 17-9. 망막의 체제(Retinal Oorganization).
⒜ 망막의 세포 체제. 광수용기는 초자방(vitreous chamber) 근처보다는 맥락막에 더 가까이 위치하고 있음에 주목하라. (LM×290)
⒝ 수평절단면의 도해 그림에서의 시신경원반(optic disc).
⒞ 눈의 동공을 통해서 보이는 망막의 사진.
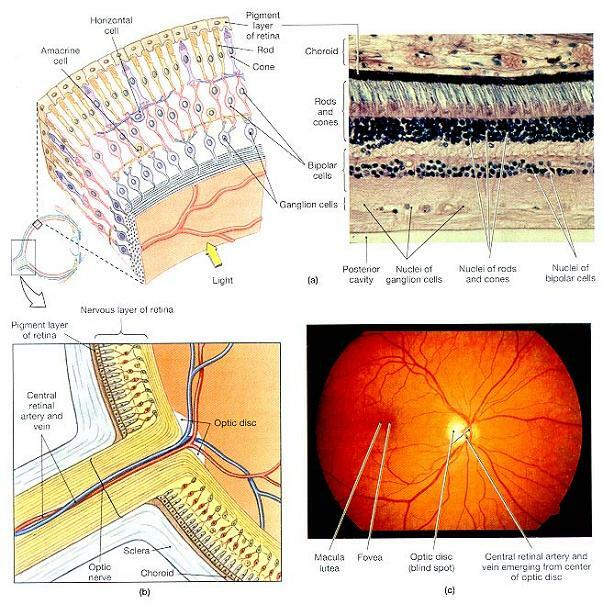
신경막(neural tunic), 즉 망막(retina)은 바깥쪽의 얇은 색소층(pigment layer)과 시각수용기 및 연계된 신경원들을 수용하고 있는 두터운 안쪽 층인 신경망막(neural retina)으로 구성되어 있다. 색소층은 신경망막을 통과한 빛을 흡수하며, 색소세포는 망막의 광수용기와 중요한 생화학적 상호작용을 한다. 신경망막은 ⑴ 빛에 반응하는 광수용기, ⑵ 시각정보의 예비 처리와 통합을 수행하는 신경원들 및 지지세포들, ⑶ 후강 내면에 배열된 조직들을 자양하는 혈관들을 내포하고 있다.
망막의 두 층은 보통 밀착하고 있지만, 단단히 연결되어 있지는 않다. 신경망막은 앞으로 단지 거상연까지만 뻗어 있지만, 망막의 색소층은 모양체와 홍채 위로 계속된다. 이렇게 해서 신경망막은 후강의 뒤와 외측 경계를 확립하는 컵을 형성한다.
망막의 체제(Retinal Organization)
단면을 보면, 망막은 여러 개의 세포층을 수용하고 있다. (그림 17-9a) 색소층에 가장 근접한 가장 바깥층이 광수용기들을 수용하고 있다.
광수용기들은 맥락막의 혈관으로부터의 산소와 양분의 확산에 전적으로 의존하고 있다. 망막박리(detached retina)에서는, 신경망막이 색소층과 분리되기 시작한다. 이 질환은 눈에 가해진 갑작스러운 충격이나 기타 여러 가지 인자들에서 초래될 수 있다. 신경막의 두 층이 재부착되지 않는다면, 광수용기들이 변성되어 시력을 상실하게 된다. 재부착은 대개 각막을 통해 초점이 맞춰진 레이저광선을 이용해 두 층을 서로 “용접하는 것(welding)”으로 시술된다.
광수용기는 막대(rods)와 원뿔(cones) 두 가지 유형이 있다. 막대는 빛의 색을 분별하지 못한다. 이들은 매우 빛에 민감해 어두운 조명의 방안이나 땅거미가 진 후 또는 희미한 달빛에서도 볼 수 있게 해준다. 원뿔은 우리에게 색이 있는 시각을 제공해준다.
원뿔에는 세 가지 유형이 있으며, 이들이 다양한 조합으로 자극되어 색들을 구별하여 지각할 수 있게 해준다. 원뿔은 더 뚜렷하고 분명한 이미지를 제공하지만, 이들은 막대보다 훨씬 강렬한 빛을 필요로 한다. 만일 여러분이 해질 무렵 야외에 앉아 있다면, 여러분의 시각 시스템이 원뿔 토대 시각(총천연색의 명확한 이미지)에서 막대 토대 시각(흑백의 비교적 점묘화같은 이미지)으로 이동하는 때를 말할 수 있을 것이다. - 포유류에는 원추세포와 간상세포가 다 있으나 조류는 주로 원추세포로 되어있다. 간상세포는 1억개, 원추세포는 5백만개 정도
 막대와 원뿔은 망막의 외면에 고르게 분포되어 있지 않다. 약 1억 2,500만의 막대는 망막 둘레에 넓은 띠를 형성한다. 뒤쪽 망막면은 대략 600만 개의 원뿔이 우위를 차지하며 존재하고 있다. 이들의 대부분은 시각 이미지가 각막과 수정체를 통과한 후 도달하는 영역에 집중되어 있다. 황반(macula lutea)이라고 하는 이 영역에는 원뿔은 존재하지 않는다. 원뿔은 중심와(fovea centralis) 또는 와(fovea)라 부르는 황반의 중심부에 가장 집중되어 있다.(맹금류는 중심와에서의 원추세포밀도가 인간의 3배이상) 이 와는 가장 뚜렷한 시력을 보이는 부위로, 여러분이 어떤 물체를 직시할 때 그 이미지는 망막의 이 부위에 맺힌다. (그림 17-8c) 물체의 중심에서 시작해 수정체 중심을 지나 중심와까지 이은 선을 눈의 시축(visual axis)으로 삼는다.
막대와 원뿔은 망막의 외면에 고르게 분포되어 있지 않다. 약 1억 2,500만의 막대는 망막 둘레에 넓은 띠를 형성한다. 뒤쪽 망막면은 대략 600만 개의 원뿔이 우위를 차지하며 존재하고 있다. 이들의 대부분은 시각 이미지가 각막과 수정체를 통과한 후 도달하는 영역에 집중되어 있다. 황반(macula lutea)이라고 하는 이 영역에는 원뿔은 존재하지 않는다. 원뿔은 중심와(fovea centralis) 또는 와(fovea)라 부르는 황반의 중심부에 가장 집중되어 있다.(맹금류는 중심와에서의 원추세포밀도가 인간의 3배이상) 이 와는 가장 뚜렷한 시력을 보이는 부위로, 여러분이 어떤 물체를 직시할 때 그 이미지는 망막의 이 부위에 맺힌다. (그림 17-8c) 물체의 중심에서 시작해 수정체 중심을 지나 중심와까지 이은 선을 눈의 시축(visual axis)으로 삼는다.
여러분은 아마도 이런 분포의 시각적 중요성을 이미 인식하고 있을 것이다. 여러분이 무언가를 직시하고 있다면, 여러분은 색채 시각의 중심인 중심와에 상이 맺히게 하고 있는 것이다. 원뿔을 충분히 자극할 수 있는 빛이 있다면, 여러분은 매우 좋은 이미지를 얻을 수 있다. 매우 어스레한 빛에서는 원뿔은 전혀 기능하지 못한다. 예를 들면, 여러분이 어슴프레한 별을 응시하려고 애쓴다면, 별을 볼 수가 없다. 그러나 만일 별을 똑바로 보지 않고 약간 옆으로 비껴 본다면 매우 선명하게 볼 수 있을 것이다. 여러분이 시선을 이동하면 별의 상이 중심와에서 주위로 이동하게 되며, 별이 제공하는 빛은 원뿔을 자극하기에는 충분치 못하지만 훨씬 민감한 막대에는 영향을 미칠 수 있기 때문이다.
막대와 원뿔은 대략 600만 개의 양극세포(bipolar cells)와 시냅스한다.(그림 17-9) 다시 양극세포들은 후강과 인접한 신경절세포(ganglion cells)로 된 층 안에서 시냅스한다. 수평세포(horizontal cells) 망은 광수용기와 양극세포 사이의 시냅스 수준에서 망막의 바깥부분을 질러 뻗어 있다. 이에 필적할만 한 아마크린세포(amacrine cells)로 된 층은 양극세포가 신경절세포와 시냅스하는 곳에서 생긴다. 수평세포와 아마크린세포는 광수용기와 신경절세포 사이의 연락을 촉진하거나 억제하여 망막의 감수성을 조정한다. 그 효과는 텔레비젼에 설정된 “콘트라스트(contrast)” 조정에 비견할 수 있다. 이들의 활동은 어두침침한 환경이나 밝은 환경에 맞춰 눈을 조정하는데 중요한 역할을 한다. - 측방향억제

그림 17-10. 시신경원반
(The Optic Disc).
왼쪽 눈을 감고 오른쪽 눈으로 십자 표시를 시야 중심에 오도록 하고 응시하라. 이 면을 몇 인치 떨어지게 한 상태에서 시작하여 점차 멀어지게 한다. 점의 상이 맹점에 도달하면 점이 보이지 않게 된다. 왼쪽 눈의 맹점을 확인하려면, 오른쪽 눈을 감고 왼쪽 눈으로 이번에는 십자 표시가 아니라 점을 응시하면서 이 과정을 되풀이한다.
시신경원반(The Optic Disc; 시신경유두)
약 100만 개로 추정되는 신경절세포로부터 나온 축색들은 중심와의 바로 외측의 둥근 영역인 시신경원반(optic disc)으로 모인다. 이 시신경원반은 시신경(NⅡ)의 기시점이다. 이 지점으로부터, 축색들은 안구의 벽을 관통해 간뇌로 진행한다.(그림 17-9b) 망막에 공급되는 중심망막동맥(central retinal artery)과 중심망막정맥(central retinal vein)은 시신경의 중심을 통과해 시신경원반의 표면에서 빠져나간다.(그림 17-9b, c) 시신경원반에는 광수용기나 기타 망막구조물이 없다. 이 부위에 빛이 도달하더라도 알아채지 못하기 때문에, 이를 보통 맹점(blind spot)이라고 부른다. 여러분은 시야에서 맹점을 인식하지 못하는데, 이는 주로 불수의적 안구 움직임이 시각 이미지를 이동시키며 뇌로 하여금 놓친 정보를 채우게 하기 때문이다. 그림 17-10은 시야에서 맹점의 존재와 위치를 입증해줄 것이다.
3.시각 생리(Visual Physiology)

그림 17-16. 막대세포와 원뿔세포(Rods and Cones).
망막의 간상세포(막대세포,rods)와 원추(원뿔세포,cones)는 가시광선의 기본단위인 광자(photon)를 탐지하기 때문에 광수용기(photoreceptor)라 불린다. 복사에너지(radiant energy)의 한 형태인 빛은 고유의 주파수(frequency)와 파장(wavelength)을 지닌 파동으로 방사된다. 가시광선은 장파인 라디오선에서부터 단파인 감마선에 이르는 전자기파 복사의 전영역 중에서 작은 일부분에 불과하다.
우리의 눈은 가시광선 영역, 즉 400~700 ㎚의 파장에 민감하다. 무지개에서 보이는 이러한 영역의 스펙트럼은 ROY G. BIV(red, orange, yellow, green, blue, indigo, violet; 빨,주,노,초,파,남,보)라는 약자로 기억할 수 있다. 빨강색 빛의 광자가 가장 적은 에너지를 가지고 있으며, 보라색 영역의 광자가 가장 많은 에너지를 가지고 있다. 막대(rods)는 파장에는 관계없이 광자의 유무에 대한 정보를 중추신경계에 제공한다. 원뿔(cones)은 파장에 대한 정보를 제공하여 우리가 색을 지각할 수 있게 해준다.
막대와 원뿔의 구조(Anatomy of Rods and Cones)
그림 17-16은 막대와 원뿔의 구조를 비교하고 있다. 길다란 광수용기 외절(outer segment)은 수백에서 수천의 편평한 막성판, 즉 디스크(disc)를 수용하고 있다. 막대(rod)와 원뿔(cone)이라는 이름은 외절의 형태를 지칭하는 것이다. 막대에서, 각 디스크들은 각기 독립적인 실체(independent entity)이며, 외절은 길다란 원통을 형성한다. 원뿔에서, 디스크들은 세포막이 주름잡힌 것(infoldings of the cell membrane)이며, 외절은 점차 가늘어져 무딘 끝을 이룬다. 좁다란 연결자루(connecting stalk)가 외절을 통상적인 세포소기관 모두를 수용하고 있는 부위인 내절(inner segment)에 부착시킨다. 내절은 다른 세포들과 시냅스 접합을 하고 있으며, 그 곳에서 신경전달물질을 유리한다.
-광수용기의 disc는 세포막이 함입되어 이루어진 것, disc는 원래 세포막에서 유래됨 →
진핵생물들은 세균들의 표면에 있는 변덕스러운 거품에 다름아니다.(리처드 도킨스)
-소위 말하는 세상을 본다할 때 빛에너지를 어떻게 받아드리는가? 어떻게 brain이 빛의 쏟아지는 알맹이들로 외부의 이미지를 만들어 내는가? 외계를 묘사하는가? → 그 출발점은 원래 있던 한 세포가 우연히 빛을 획득하는 능력을 가지게 됨, 세포막이 함입되어 disc를 형성, 그 disc에 로돕신이란 단백질이 삽입되어있고 그 단백질의 한 부위가 retinal이라는 분자 구조이고 그 분자구조가 빛을 획득하는 능력을 가짐.
시각색소(Visual Pigments)
막대와 원뿔 모두에 있는 외절의 디스크는 시각색소(visual pigment)라 하는 특수한 유기화합물(organic compound)을 내포하고 있다. 시각색소에 의한 광자의 흡수가 광수용의 과정의 으뜸가는 핵심 단계이다. 시각색소는 로돕신(rhodopsin) 화합물의 유도체이다. 로돕신은 옵신(opsin)이란 단백질에 비타민A로부터 합성된 레티날(retinal)이란 색소가 결합되어 구성된다. 그림 17-16은 로돕신 분자의 구조를 도해하고 있다.
막대와 원뿔 모두에서 동일한 망막색소(retinal pigment)가 발견되지만, 옵신은 네 가지 형태가 있다. 존재하는 옵신의 유형이 레티날에 의해 흡수될 수 있는 빛의 파장을 결정한다. 모든 막대는 한 가지 형태의 옵신으로 이루어져 있다. 원뿔은 세 가지 다른 집단이 있는데, 이들은 각기 다른 형태의 옵신으로 되어 있으며, 또 각기 다른 범위의 파장을 감지한다. 원뿔 집단에 가해지는 자극의 감별이 색각(color vision)의 기초가 된다.
시각색소를 내포하고 있는 새로운 디스크들은 외절의 기저부에서 끊임없이 조립된다. 이어서 완성된 디스크는 외절의 끝을 향해 이동한다. 약 10일 후, 디스크는 작은 세포질 방울 안에 들어 떨어져 나간다. 이 방울들은 색소세포들에 의해 흡수되며, 이 세포들은 막성분을 분해하고 레티날을 비타민A로 다시 전환시킨다. 이어서 비타민A는 색소세포내에 저장되었다가 후에 광수용기로 전달된다.
광수용(Photoreception)
이제부터는 로돕신에 기초한 광수용(rhodopsin-based photoreception)의 단계들을 쫒아가기로 한다. 어둠 속에서, 광수용기 외절의 세포막은 열려져 있는 화학적으로 조절되는 나트륨이온 채널들을 수용하고 있다. 이 관문채널들은 고에너지화합물인 GTP의 유도체인 cyclic-GMP(cGMP)의 존재 하에 계속 열려져 있다. 이렇게 채널이 열려있기 때문에, 이들의 막전위는 전형적인 신경원의 휴지전위인 -70 ㎷가 아니라 약 -40 ㎷ 정도가 된다.
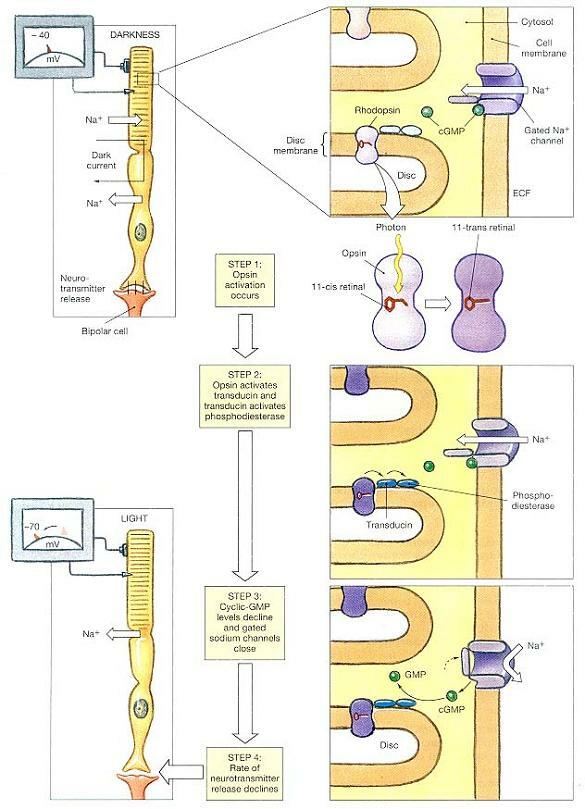
내절은 세포질 밖으로 나트륨이온을 계속해서 퍼내는데, 외절 안으로 들어와 다시 내절로 이동한 후 다시 세포 밖으로 내보내지는 나트륨이온의 이 이동을 암전류(dark current)라 한다. (그림 17-17)
이 막전위에서, 광수용기는 내절에 있는 시냅스에서 계속 신경전달물질을 유리하고 있다. 광자의 도착은 암전류를 감소시키고 막전위를 변경시켜 신경전달물질 유리속도가 변하게 한다.

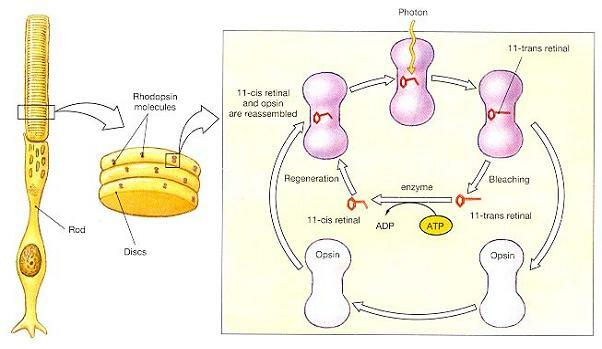
이 과정은 광자가 로돕신 분자의 레티날 부분을 때릴 때 시작된다. 결합 레티날 분자의 가능한 배열은 두 가지가 있다. 이들은 평상시에는 11-cis형으로 있다가, 빛을 흡수하면 보다 선형인 11-trans형을 취한다. 이 형태의 변화는 그림 17-17에 도해되어 있는 일련의 효소 단계들을 격발한다.
1 단계(Step 1): 옵신의 활성화가 일어난다(Opsin activation occurs).
로돕신은 디스크의 막 안에 끼워넣어져 있는 수용체단백(receptor protein)이다. 옵신은 효소이며, 어둠속에서는 활동하지 않는다. 빛이 로돕신 분자를 때리면 레티날이 11-cis형에서 11-trans형으로 전환된다. 이 전환은 로돕신 분자의 옵신 부분을 활성화 시킨다.
2 단계(Step 2): 옵신이 트랜스듀신이란 2차효소를 활성화시키고, 트랜스듀신은 포스포디에스테라제를 활성화시킨다(Opsin activates a second enzyme, called transducin, and transducin activates phosphodiesterase(PDE)).
트랜스듀신은 세포막의 수용체 단백과의 상호작용으로 활성화되는 막결합효소인 G단백질(G-protein)의 하나이다. 이 경우, 트랜스듀신은 옵신에 의해 활성화되고, 이어서 트랜스듀신은 포스포디에스테라제(PDE)를 활성화시킨다. PDE는 cyclic-GMP(cGMP)를 분해하는 효소이다.
3 단계(Step 3): cGMP 수준이 떨어지고 나트륨이온채널이 닫힌다(Cyclic-GMP levels decline, and gated sodium ion channels close).
나트륨 채널에서 cGMP가 떨어져 나가면 채널이 불활성화된다. 이에 따라 세포내로의 나트륨 이온의 유입 속도가 감소된다.
4 단계(Step 4): 신경전달물질 유리속도가 떨어진다(The rate of neurotransmitter release declines).
능동수송이 세포질내에서 나트륨이온 제거를 계속하기 때문에, 나트륨 채널이 닫히면 막전위는 -70 ㎷를 향해 떨어진다. 막전위가 떨어지면 신경전달물질 유리 속도도 떨어진다. 이런 저하는 해당 수용기가 광자를 흡수했다는 것을 나타내는 것이다.
 쌍극세포에서 글루탐산 receptor에 따라 달라진다.
쌍극세포에서 글루탐산 receptor에 따라 달라진다.
암반응 시, 로돕신 불활성으로 신경전달물질인 Glutamate released → Bipolar cell either depolarized or hyperpolarized, depending on glutamate receptor
명반을 시, 로돕신 활성으로 Na+channels closed → Rod hyperpolarized → No glutamate released → Bipolar cell either → Bipolar cell either hyperpolarized or depolarized, depending on glutamate receptor
자극후 복구(Recovery After Stimulatuion)
광자를 흡수한 후 레티날이 11-cis형으로의 재전환이 자연스럽게 일어나지는 않는다. 오히려 로돕신 분자 전체가 반드시 분해되었다가 재조립되어야만 한다. 분자구조의 변화가 일어나고 얼마 뒤, 로돕신 분자는 레티날과 옵신으로 분해되기 시작하며, 이 과정을 표백(bleaching)이라 한다. 레티날이 옵신과 재결합할 수 있으려면 반드시 효소에 의해 11-cis형으로 전환되어져야만 한다. 이 전환은 ATP 형태로 에너지를 필요로 하며, 시간도 걸린다. 표백이 사진찍을 때 번쩍인 섬광전구가 꺼진 후에도 잔상이 남게 만든다. 빛에 강하게 노출된 후, 광수용기는 로돕신 분자가 재생될 때까지 더 이상 자극에 반응하지 못한다. 그 결과, “허상(ghost image)”이 망막에 남게 된다. 표백은 평상시의 환경에서는 결코 느껴지 못하는데, 이는 눈이 상이 맺히는 망막면의 위치가 조금씩 바뀌도록 하는 불수의적 변화를 끊임없이 일으키고 있기 때문이다. 로돕신 분자가 재조립되는 동안, 막투과성은 정상으로 돌아간다. 옵신은 표백이 일어날 때 비활성화되고, 그 결과로 cGMP의 분해가 멈춘다. 다른 효소들이 cGMP를 세포질내에 생산하면, 화학적으로 개폐되는 나트륨채널이 다시 열리게 된다.
원뿔세포에는 세 가지 유형이 있다. 청색원뿔(blue cones), 녹색원뿔(green cones), 적색원뿔(red cones). 각 유형은 각기 다른 형태의 옵신과 다른 범위의 감수성을 가지고 있다. 이들이 다양한 조합으로 자극되는 것이 색의 지각을 담당한다. 정상적인 사람의 원뿔세포는 청색원뿔 16%, 녹색원뿔 10%, 적색원뿔 74%로 구성되어 있다. 비록 이들의 감수성이 서로 겹치기는 하지만, 각 유형은 각기 특정 가시광선 영역에서 가장 민감하다.
색 식별(color discrimination)은 세 유형의 원뿔세포 모두에서 온 정보의 통합을 통해 일어난다. 예를 들면, 노랑색으로의 지각은 녹색원뿔(매우 자극됨)과 적색원뿔(자극됨) 및 청색원뿔(비교적 영향받지 않았음)에서 온 입력의 조합으로 이루어진다. 만일 세 가지 원뿔들이 모두 자극되었다면, 우리는 흰색으로 지각할 것이고, 아울러 원뿔이 아니라 막대가 자극받았다면 이 역시 우리는 흰색으로 지각한다. 이러한 이유로, 어두침침한 곳으로 들어가거나 별빛에 의지해 걷고 있을 때에는 모든 물체가 검은색과 흰색으로만 보이게 된다.
명순응과 암순응(Light and Dark Adaptation)
시각 시스템의 감수성은 조도(intensity of illumination)에 따라 차이가 난다. 어둠 속에 들어가 30분 이상의 시간이 지나면, 거의 모든 시각색소들이 자극을 최대로 받아들이게 된다. 이것이 암순응 상태(dark-adapted state)이다. 암순응이 되면 시각 시스템은 극도로 민감해진다. 예를 들어, 단 하나의 광자에 반응해 막대세포 하나가 과분극된다. 더욱 주목할 만한 것은, 만약 7개 정도의 막대들이 동시에 광자를 흡수한다면, 그 사람은 섬광을 보게 된다.
빛이 들어오면, 처음에는 거의 견딜 수 없을 정도로 밝지만, 몇 분이 지나면 표백(bleaching)이 일어나면서 감수성이 낮아지게 되고, 결국 시각색소의 분해 속도와 재형성 속도가 균형을 이루게 된다. 이 상태가 명순응 상태(light-adapted state)이다. 사람이 어두운 동굴에서 밝은 대낮으로 나올 때, 수용기 감수성은 약 25,000 배 감소한다.
여러 가지 중추성 반응들이 광감수성을 더욱 더 조정해준다. 동공축소반사(pupillary constrictor reflex)를 통한 동공의 축소는 눈으로 들어오는 빛의 양을 최대 암순응 수준의 1/30로 줄인다. 동공이 최대로 확대되는 동안에는, 눈으로 들어오는 빛의 양이 30 배 증가되며, 시각로상의 일부 시냅스들의 촉진은 감수성을 3 배 정도 상승시킬 수 있으며, 시스템 전체가 한 가지 인자에 의해 100만 배 이상 그 효율성이 상승하기도 한다.
4.시각로(The Visual Pathway)
시각로는 광수용체기서 시작해 대뇌반구의 시각피질에서 끝난다. 우리가 검토한 바 있는 다른 감각 경로들에서는, 정보를 중추신경계로 보내는 감각신경원과 수용기 사이에는 기껏해야 하나의 시냅스가 있다. 시각로에서, 모든 메시지는 반드시 두 개의 시냅스(광수용기-양극세포, 양극세포-신경절세포)를 지나야만 뇌로 향할 수 있다. 이 여분의 시냅스는 시냅스 지연이 더해지게 하지만, 시각정보가 망막을 떠나기 전에 처리되고 통합될 수 있는 기회를 제공한다.
망막 처리(Retinal Processing) - receptive field
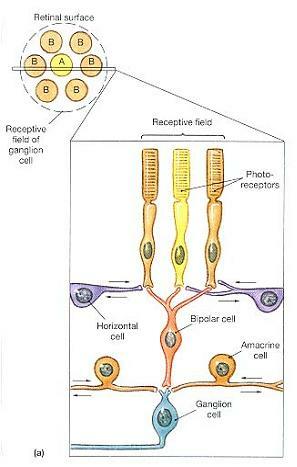
망막에 있는 각각의 광수용기는 각기 특정 수용야(specific receptive field)를 모니터한다.
그림 17-20. 수렴과 신경절세포의 기능(Convergence &
Ganglion Cell Function).
⒜ 신경절세포는 영역이 뚜렷한 시야 부분을 모니터한다.
⒝ 어떤 신경절세포들은 수용야(receptive field) 중앙에 도달하는 빛에 강하게 반응한다
(on-center neurons). 다른 신경절세포들은 수용야 가장자리의 조도에 가장 강하게 반응한다
(off-center neurons).
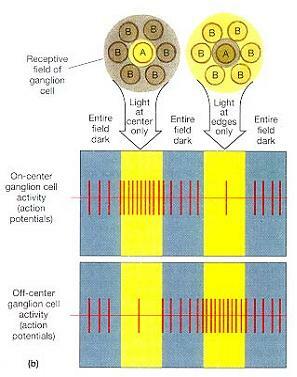
망막은 약 1억 3,000만 개의 광수용기와 600만 개의 양극세포 및 100만 개의 신경절세포들을 수용하고 있다. 따라서 시각로의 시작점에는 상당한 양의 수렴(convergence)이 있게 된다. 수렴의 정도는 여러분이 막대를 고려하냐 원뿔을 고려하냐에 따라 차이가 난다. 수렴의 양에 관계없이 각각의 신경절세포들은 시야(visual field)의 특정 부분들을 모니터한다.
약 1,000 개 정도의 막대들이 자신들의 양극세포들을 경유해 하나의 신경절세포로 정보를 보낸다. 막대들을 모니터하고 있는 신경절세포들은 비교적 크며, 이들을 M 세포(M cells; magno-cell)라 부른다. M 세포들은 어두침침한 빛의 상태에서 전반적인 외형, 동작 및 음영에 대한 정보를 제공한다. 매우 많은 수렴을 하기 때문에, M 세포 하나가 활동하기 시작했다면 이는 빛이 어떤 특정 위치가 아니라 전반적인 영역에 도달했다는 것을 의미한다.
수렴으로 기인된 국한성(specificity)의 상실은 신경절세포들은 자신의 감각야(sensory field)에서의 활동 패턴에 따라 활동을 바꾼다는 사실에 의해 일부 극복된다. 감각야는 대부분 둥글며(그림 17-20a), 신경절세포들은 전형적으로 자신들의 수용야의 중앙에 도달한 자극과 가장자리에 도달한 자극에 다르게 반응한다. 어떤 신경절세포들은 감각야의 중앙에 도달한 빛에 의해 흥분되고(중심형 신경원(on-center neurons)), 빛이 수용야의 가장자리를 때릴 때에는 억제된다. 이와 다른 신경절세포들은 중심부를 비추는 빛에 의해 억제되고 가장자리를 비추는 빛에 의해 자극된다(주변형 신경원(off-center neurons)). 이 둘의 관계는 그림 17-20b에 도해되어 있다. 중심형(on-center)과 주변형(off-center) 신경원들은 그들의 수용야의 어떤 부분에 빛이 비쳐지는지에 대한 정보를 제공한다.
뿔들에서는 전형적으로 수렴이 매우 조금밖에 일어나지 않으며, 중심와에서 원뿔세포와 신경절세포의 비율은 1:1이다. 원뿔들을 모니터하는 P 세포(P cells; parvo cells)라 하는 신경절세포들은 M 세포보다 크기는 작고 숫적으로는 더 많다. P세포는 밝은 빛에서 활동하며, 이들은 가장자리의 매우 상세하게 색에 대한 정보를 제공한다. 수렴이 별로 일어나지 않기 때문에, P 세포 하나의 활성화는 하나의 특정 위치로 빛이 도달했다는 것을 의미한다. 그 결과, 원뿔은 시각 이미지에 대한 보다 정확한 정보를 제공한다. 사진학적 관점에서 볼 때, 막대들에 의해 형성된 사진은 세부사항이 흐릿한 조잡하고 입자가 거친 모습을 보이며, 반면에 원뿔들에 의해 생성된 사진은 입자가 곱고 선명하며 뚜렷하다
 -측방향억제
-측방향억제
-외측슬상체의 6개층중 1,2.층은 M세포로 움직임을 감지, 3~6층은 P세포로 형태,색을 process를, 망막에서의 6개층이 LGN과 VI까지 그대로 mapping 된다.
-반대측 눈에서 정보가 LGN의 1,4,6층으로
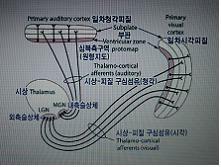
외측, 내측슬상체의 구조가 대뇌 피질(1차영역)까지 map형태로 그대로 따라간다.
시각정보의 중추 처리(Central Process ing of Visual Information)
ing of Visual Information)
신경절세포들 전체에서 나온 축색들은 시신경원반(optic disc)에 집중해, 눈의 벽을 관통한 뒤, 시신경(N Ⅱ)으로서 간뇌(diencephalon)를 향해 나간다. 양쪽 눈에서 하나씩 나오는 두 시신경은 시신경 교차(optic chiasm)에서 간뇌에 도착한다. (그림 17-21). 이 지점에서 신경섬유들의 약 절반 정도는 같은 쪽 뇌의 외측슬상체(lateral geniculate, 2,3,5층)를 향해 나가고, 나머지 절반은 교차하여 반대쪽 외측슬상체(1,4,6층)로 간다. 외측슬상체는 대뇌피질 뿐만 아니라 뇌간의 반사중추로도 시각정보를 중계하는 개폐 및 처리 중추(switching and processing center)로 작용한다. 예를 들어, 동공반사와 안구 움직임을 제어하는 반사들은 외측슬상체에 의해 상구(superior colliculus)로 중계되는 정보에 의해 격발된다.
각 외측슬상체에서 나온 시각정보는 같은 쪽 대뇌반구의 후두피질로 간다. 외측슬상체와 시각피질을 연결하는 투사섬유 다발은 시각로부채살(optic radiation)이라 한다.
그림 17-21. 시각로(The Visual Pathway).
시신경교차에서 신경섬유들의 일부 교차가 일어난다. 그 결과 각 반구는 같은 쪽 눈의 시야의 외측 절반에서 온 시각정보와 반대쪽 눈의 시야의 내측 절반에서 온 시각정보를 받아들이게 된다. 시각연합영역은 이 정보를 통합해 전체 시야의 완전한 사진을 이루어낸다. - 개구리의 경우 거의 대부분 교차된다
시야(The visual field)
시각 이미지의 지각은 도달한 정보의 후두엽의 시각 피질에서의 통합을 반영한다. 각각의 눈은 다음과 같은 이유로 약간 다른 시각 이미지를 받아들인다. ⑴ 양 눈의 중심와(fovea)는 5~7.5 cm 떨어져 있으며, ⑵ 코와 안와가 반대쪽 시계를 보지 못하도록 차단하고 있다. 깊이 지각(depth perception), 즉 시계내 대상체들 사이의 삼차원적 관계의 해석은 왼쪽 눈과 오른쪽 눈이 받아들이는 이미지 안에서의 상대적인 위치를 비교하는 것으로 얻어진다.
여러분이 앞을 보고 있다면, 왼쪽 눈과 오른쪽 눈에서 들어오는 시각 이미지는 그림 17-21에 나타내진 것처럼 겹쳐진다. 각 눈의 중심와에 의해 받아들여지는 이미지는 겹쳐지는 부위의 중심에 위치한다. 겹쳐지는 부위의 중심을 지나도록 그은 수직선이 시신경교차(optic chiasm)에서의 시각정보 분할(division of visual information)의 경계를 정한다. 왼쪽 절반의 결합 시야에서 오는 시각정보는 왼쪽 후두엽의 시각피질에 도달할 것이며, 오른쪽 절반의 결합 시야에서 오는 시각정보는 왼쪽 시각피질에 도달할 것이다.(Visual information from the left half of the combined visual field will reach the visual cortex of the left occipital lobe; visual information from the right half of the combined visual field will arrive at the left visual cortex).
그리하여 대뇌반구들은 전체 시야의 지도 하나를 담게 된다. 1차감각피질의 경우에서와 같이, 그 지도는 감각야 안의 상대적인 부위들을 똑같은 비율로 배분하지 않고 있다. 예를 들면, 황반과 중심와에 할당된 영역은 정비례할 때 배당되는 면적의 약 35 배 정도를 차지하고 있다. 또 그 지도는 망막에서의 시각 이미지의 방향과 똑같이 상하와 전후가 바뀌어 있다.
-영장류의 후뇌 피질의 60%는 시각을 process 한다.
-VI에서 두정엽PG로 가는 흐름 : 움직임, 위치등을 process
/ 측두엽TE로 가는 흐름 : 형태, 모양, 색깔을 process
-VI영역에서 단순세포(선과 점을 감지), 복합세포(선의 이동을), 초복합세포(모서리 가진 부분)로 나뉜다.
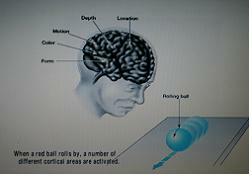
시각은 여러부위에서 process 되고 있다.
굴러가는 공을 볼 때 대뇌피질 여러부위가 같이 활동, 형태, 색깔은 측두엽에서 / 움직임, 깊이, 위치는 두정엽에서 등등..
5.뇌간과 시각 처리(The Brain Stem and Visual Processing)
뇌간내의 많은 중추들은 외측슬상체로부터 또는 시삭의 부축색(collaterals)으로부터 시각정보를 받아들인다. 외측슬상체를 그냥 지나치는 부축색들은 상구나 시상하부에서 시냅스한다. 중뇌의 상구는 시각 자극에 반응해 눈과 머리 및 목의 무의식적 움직임을 제어하는 운동명령을 내보낸다. 시상하부의 교차상핵(suprachiasmatic nucleus)으로의 시각정보 입력은 뇌간의 다른 핵들의 기능에 영향을 미친다. 이 핵과 시상상부의 송과체(pineal gland)가 낮/밤 주기(day/night cycle)와 결부되어 있는 내장활동의 일상 패턴을 확립한다. 이 日周期 리듬(circadian rhythm)은 대사율, 내분비 기능, 혈압, 소화 활동, 각성/수면 주기와 또 14~16 장에서 논의된 바 있는 기타 생리적 및 행동적 과정에 영향을 미친다.
6.눈의 진화

진화론적으로
1)바늘구멍눈 ex)앵무조개
2)거울눈 -반사층에 반사되어 망막에 상이 맺힘 ex)조개류
3)카메라눈 - 렌즈(수정체)가 발생
-피부세포가 시각세포로 되는데 1세대에 폭과 길이, 단백질 농도 등이 1% 정도만 변화해도 40만세대(50만년)가 지나면 지금과 같은 카메라눈이 발생가능하다는 시뮬레이션실험이 있었다
- 최근 복잡한 연구에 의해도 100만년이면 카메라눈이 발생 가능
-캄브리아기 생명의 대폭발(5억4천3백만년전) 보다 앞서 지금으로부터 5억4천4백만년전에 눈이 생김
-신생아 VI의 신경섬유 연결이 성긴데 6개월 후면 성인의 수준이 된다.
-“눈의 탄생“의 저자, 앤드루 파커에 의하면 캄브리아기 생명의 대폭발의 첫 스타트가 눈스위치 이론이다. 어떤 동물이 눈을 획득하게 되면서 무기의 무한 경쟁이 시작되었다. 판피류, 악구류 등등 생겨남. 청각, 촉각은 발신자와 수신자가 따로 있으나 시각은 수신자만 존재한다. 빛에 대해선 그 영향 내에 다 드러나 있기에 대처하지 않을 수 없다.→ 위장camouflage
7.Receptive Fields
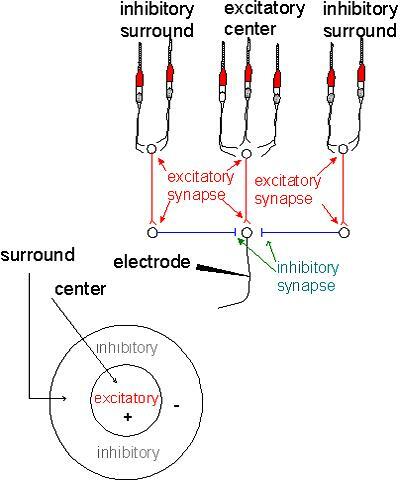
그림의 위쪽은 광수용기이고, 아래쪽은 두 종류의 시냅스를 보여준다.
Excitation synapses는 신경 활동성을 증가시키고, inhibitory synapses는 감소시킨다.
동심원은 수용기가 빛에 의해 자극받을 때,
electrode전극으로 표시되는 neural activity를 나타낸다.
center 수용기의 하나 내지는 모두가 자극될 때
neural activity의 excitatory increase가 electrode에 획득되어진다. surround 수용기들이 자극되면 neural activity의 inhibitory decrease가 electrode에 획득되어진다.